Как изготавливается сахар из сахарной свеклы
Поставка сахарной свеклы. После сбора урожая свеклу транспортируют на сахарный завод.
Выборочный контроль. После транспортировки свекла проходит выборочную проверку. Сумма оплаты плодоводу зависит от количества чистых плодов и содержания в них сахара.
Очистка. Свеклу взвешивают и делят на большие партии. Оттуда овощи транспортируются в устройство для промывки свеклы, где овощи фильтруют от камней и песка.
Обрезка. Чтобы извлечь сахар, свеклу измельчают до состояния стружки – тонких полосок наподобие картофеля-фри.
Диффузия. Для извлечения сахара из свеклы стружка проходит через воду, подогретую до температуры 70°C. Мякоть, которая остается после извлечения сахара (свекловичный жом), используется для изготовления корма для животных и других продуктов.
Очистка сахарного сиропа. Теплый сахарный сироп (сырой сок) содержит около 15% сахара, а также 1-2% примесей (не сахарных), которые должны быть устранены. Их удаление выполняется с помощью оксида кальция (жженой извести).
Очищенный сок становится густым. Сахарный сок, который на этом этапе уже являет собой бледно-желтую жидкость густой консистенции, называется очищенным соком. Этот сок помещается в установку для термического выпаривания с целью устранения воды, в результате чего сок становится еще более густым. Конечная жидкость, называемая густым соком, содержит приблизительно 70% сахара.
Кристаллизация. Густой сок под давлением подается в большие варочно-кристаллизационные станции, в которых формируются крошечные сахарные кристаллики.
Центрифугальное формирование.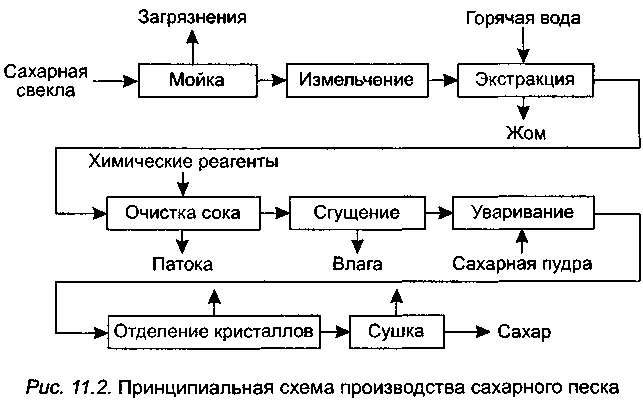 Густой коричневый сок, который на этом этапе называется утфелем, вращается в центрифуге для отделения белого сахара от коричневого сиропа. Сироп возвращается в варочно-кристаллизационную станцию и поддается повторному кипячению до момента, пока весь сахар не будет извлечен из сырья.
Густой коричневый сок, который на этом этапе называется утфелем, вращается в центрифуге для отделения белого сахара от коричневого сиропа. Сироп возвращается в варочно-кристаллизационную станцию и поддается повторному кипячению до момента, пока весь сахар не будет извлечен из сырья.
Меласса. Оставшийся продукт носит название «меласса». Содержание в нем сахара слишком низок, чтобы использовать его для выработки сахара. Меласса используется для изготовления кормов для животных, дрожжей и спирта.
Готов к использованию. В конце сахар высушивают и помещают для хранения в силосные ямы. По истечении сезона производства сахарной свеклы силосные ямы заполнены полностью, но в течение года они будут опустошаться по мере сбыта сахара в магазины, промышленные предприятия или с целью экспорта.
ТЕРМИНЫ, ИСПОЛЬЗУЕМЫЕ В ТЕХНОЛОГИИ САХАРА
Адсорбционная очистка сахаросодержащих растворов — удаление несахаров из сахаросодержащих растворов адсорбентами
Аффинация сахара — повышение чистоты сахара путем замены несахаров в пленке межкристального раствора на поверхности кристаллов посредством их диффузии в сахаросодержащий раствор более высокой чистоты при их смешивании
Бестарное хранение сахара – хранение сахара насыпью в складах силосного или ангарного типа.
Брак свекловичной стружки – показатель, характеризующий содержание в массе свекловичной стружки мелких кусочков свекловичной стружки, мезги, а также неразрезанных пластинок, не отвечающих установленным показателям качества свекловичной стружки, выраженный в процентах к массе сахарной свеклы.
Вентилирование кагатов сахарной свеклы – принудительный воздухообмен в кагатах сахарной свеклы с целью поддержания оптимальных условий хранения.
Вторая дефекация – стадия известково-углекислотной очистки, на которой осуществляется взаимодействие фильтрованного сока первой ступени сатурации с известковым молоком с целью разложения несахаров.
Вторая ступень сатурации – стадия известково-углекислотной очистки, на которой сок после второй дефекации взаимодействует с диоксидом углерода до достижения заданного значения рН и щелочности.
Гашение извести – взаимодействие извести с водой в целях превращения оксида кальция в гидроксид кальция при получении известкового молока.
Гидроподача сахарной свеклы – подача корнеплодов сахарной свеклы потоком воды по гидротранспортеру.
Гранулирование сушеного жома – формование гранул определенной формы и размеров из сушеного рассыпного жома в прессе-грануляторе.
Двухступенчатая кристаллизация сахарозы – совокупность технологических процессов и операций, объединяющих две последовательные стадии кристаллизации сахаросодержащего раствора с получением сахаров I и II кристаллизации.
Дефекация сахаросодержащего раствора – взаимодействие сахаросодержащего раствора с известковым молоком с целью разложения несахаров.
Дефекованный сок – диффузионный сок, обработанный известковым молоком.
Диффузионный сок – сок, извлеченный из свекловичной стружки в процессе экстрагирования сахарозы.
Жом – свекловичная стружка, обессахаренная в диффузионном аппарате.
Жомопрессовая вода – вода, отделенная от жома в процессе его прессования.
Жом глубокого отжатия – жом, обезвоженный путем прессования до содержания сухих веществ не менее 25 %.
Жом сушеный в гранулах – сушеный жом в виде гранул определенной формы и размеров, полученных в прессе-грануляторе.
Жом сушеный в рассыпном виде – сушеный жом в виде однородной сыпучей массы.
Заводка кристаллов – стадия уваривания утфеля в вакуум-аппарате, на которой осуществляется принудительное введение центров кристаллизации сахарозы в виде затравочного материала в пересыщенный сахаросодержащий раствор
Загрязненность сахарной свеклы – показатель, характеризующий содержание в партии корнеплодов сахарной свеклы примесей органического и минерального происхождения, выраженный в процентах к массе сахарной свеклы.
П р и м е ч а н и е – К примесям органического происхождения относятся: зеленая масса, корнеплоды столовой и кормовой свеклы, головки, боковые корешки и хвостики корне-плодов сахарной свеклы; к примесям минерального происхождения – почва, камни и другие твердые примеси.
Известково-углекислотная очистка диффузионного сока – совокупность технологических процессов и операций удаления несахаров из диффузионного сока с применением в качестве реагентов извести и диоксида углерода.
Известковое молоко – продукт гашения извести, представляющий собой водную суспензию гидроксида кальция заданной плотности.
Известняковый камень – горная порода осадочного происхождения, содержащая карбонат кальция, служащая для получения извести и сатурационного газа путем обжига в известково-газовых печах.
Известь – оксид кальция с примесями, присущими известняковому камню, полученный при его обжиге.
Изрезывание сахарной свеклы – измельчение корнеплодов сахарной свеклы в свекловичную стружку.
Ингибирование накипеобразования – предотвращение отложения накипи на поверхности нагрева теплообменного оборудования технологической линии производства сахара.
Кислый жом – жом, подвергшийся брожению под действием микроорганизмов в жомохранилище.
Клерование сахара – растворение кристаллов сахара в сиропе, промое, очищенном соке, воде.
Клеровка – раствор сахара в очищенном диффузионном соке, сиропе, оттеке, промое или воде.
Корнеплод сахарной свеклы – главный корень растения сахарной свеклы, образовавшийся в первый год вегетации и используемый для производства сахара.
Красящие вещества сахарного производства – окрашенные сложные органические соединения различной степени конденсации и полимеризации, возникшие за счет химического взаимодействия в полуфабрикатах некоторых извлеченных вместе с сахарозой нативных веществ сырья между собой и с образовавшимися в ходе превращений в технологическом потоке, придающие окраску от светло-желтого до темно-коричневого цвета полуфабрикатам и готовой продукции сахарного производства.
Кристаллизация сахарозы – выделение сахарозы в виде кристаллов из пересыщенных сахаросодержащих растворов.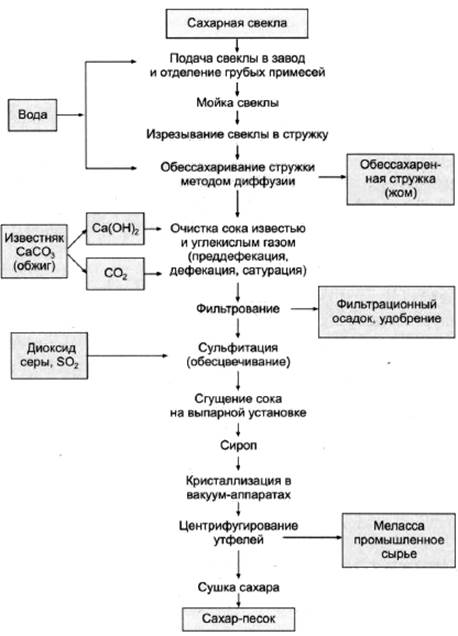
Маточный утфель – утфель, содержащий определенное количество кристаллов заданных размеров, используемый в качестве кристаллической основы для уваривания утфелей I, II и III кристаллизации.
Межкристальный раствор утфеля – жидкая фаза утфеля, представляющая собой насыщенный или пересыщенный раствор сахарозы, содержащий несахара.
Мезга – мельчайшие частицы свекловичной стружки.
Меласса – побочный продукт сахарного производства, представляющий собой межкристальный раствор, отделяемый при центрифугировании утфеля последней кристаллизации.
Мойка сахарной свеклы – удаление прилипших примесей с поверхности корнеплодов сахарной свеклы водой.
П р и м е ч а н и е – К прилипшим примесям относят землю, песок, сорняки, солому и др.
Накипеобразование – отложение накипи на поверхности нагрева теплообменного оборудования технологической линии производства сахара.
Накипь – твердые отложения, образующиеся на внутренней поверхности теплообменного оборудования при кипении сахаросодержащих растворов.
Несахара – сухие вещества, кроме сахарозы, в продуктах сахарного производства.
Обжиг известнякового камня – термическое воздействие на известняковый камень с целью разложения содержащегося в нем карбоната кальция на оксид кальция и диоксид углерода в известково-газовой печи при заданной температуре.
Обессахаривание фильтрационного осадка – извлечение сахаросодержащего раствора из пор фильтрационного осадка.
Обесцвечивание сахаросодержащего раствора – удаление красящих веществ из сахаросодержащего раствора, приводящее к снижению его цветности.
Основная дефекация – стадия известково-углекислотной очистки, на которой осуществляется взаимодействие преддефекованного сока с известковым молоком с целью разложения несахаров и формирования избытка гидроксида кальция.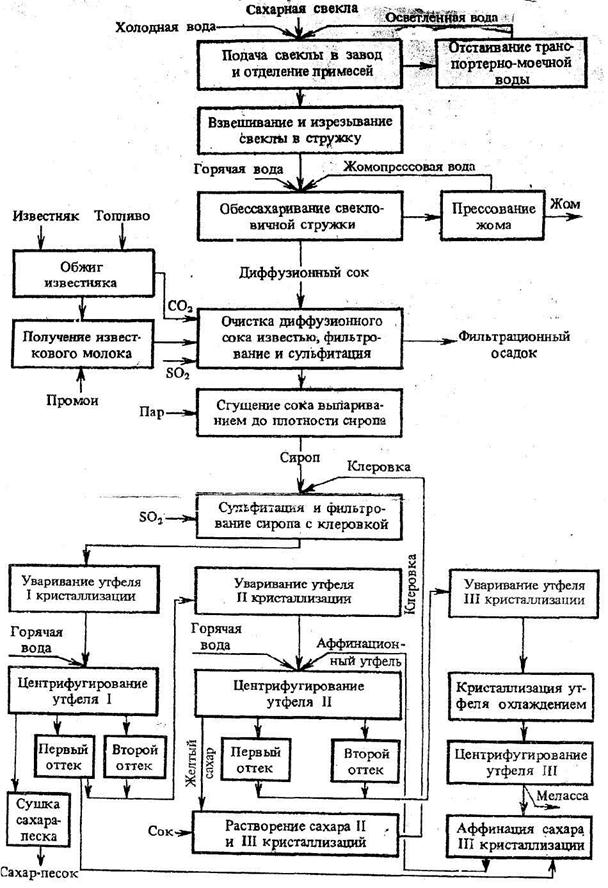
Отбор диффузионного сока – показатель, характеризующий количество отбираемого из диффузионной установки диффузионного сока, выраженный в процентах к массе сахарной свеклы.
Отжатие жома – механическое обезвоживание сырого жома.
Оттек – межкристальный раствор, отделяемый при центрифугировании утфеля.
Очистка печного газа – удаление из печного газа механических примесей и смолистых веществ с последующим его охлаждением.
Очистка сахаросодержащих растворов – совокупность технологических процессов и операций удаления несахаров из сахаросодержащих растворов с целью повышения их чистоты.
Очищенный диффузионный сок – диффузионный сок, прошедший все стадии очистки согласно технологической схеме.
Ошпаривание свекловичной стружки – тепловая обработка свекловичной стружки с целью денатурации протоплазмы клеток свекловичной ткани.
П р и м е ч а н и е – Денатурация протоплазмы клеток свекловичной ткани – необратимое изменение естественных свойств протоплазмы клеток свеклы под воздействием различных факторов.
Пеногашение – предотвращение образования или разрушение пены в сахаросодержащих растворах технологического потока производства сахара путем применения пеногасителей.
Пенообразование – образование в сахаросодержащих растворах и технологических водах технологического потока производства сахара пены, обусловленное присутствием несахаров с поверхностно-активными свойствами или жизнедеятельностью микроорганизмов.
Первая ступень сатурации – стадия известково-углекислотной очистки, на которой сок после основной дефекации взаимодействует с диоксидом углерода до достижения заданного значения рН и щелочности.
Печной газ – газ, образующийся в процессе обжига известнякового камня.
Подача сахарной свеклы – перемещение корнеплодов сахарной свеклы с кагатного поля или сплавной площадки в переработку; осуществляется способами: гидроподачи, с использованием механизмов и комбинированным.
Предварительная дефекация диффузионного сока – стадия известково-углекислотной очистки, на которой осуществляется взаимодействие диффузионного сока с известковым молоком и кальцийсодержащими реагентами с целью коагуляции высокомолекулярных соединений и осаждения нерастворимых солей кальция.
П р и м е ч а н и е – К кальцийсодержащим реагентам относятся: возращаемые суспензия сока первой ступени сатурации и суспензия сока второй ступени сатурации.
Преддефекованный сок – диффузионный сок, прошедший стадию преддефекации.
Прессование сахара – формование кусочков сахара определенной формы и размеров из увлажненного кристаллического сахара.
Производство сахара из тростникового сахара-сырца – сахарное производство, в котором используется вторичное сахароносное растительное сырье.
Промой – раствор, образующийся при обессахаривании отфильтрованного осадка или адсорбента путем промывания его водой.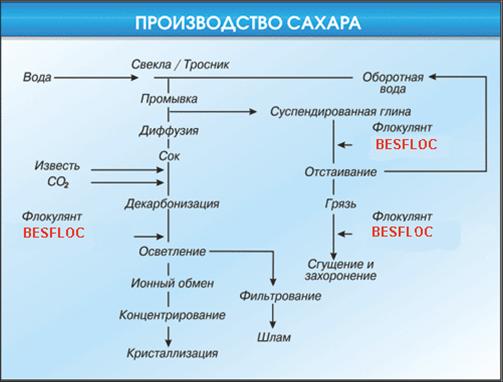
Промывание кристаллов сахарозы – удаление пленки межкристального раствора с поверхности кристаллов сахарозы в роторе центрифуги водой, сахаросодержащим раствором или паром.
Рассеивание сахара – разделение кристаллов сахара по фракциям с определенным размером.
Редуцирующие вещества сахарного производства – моносахариды сырья, готовой продукции, полуфабрикатов сахарного производства, имеющие в своем составе карбонильную группу и проявляющие свойства восстановителя в окислительно-восстановительных реакциях.
Рециркуляция сока в сатураторе – Многократное полное или частичное возвращение выходящего потока сока в аппарат.
Сатурационный газ – печной газ, содержащий диоксид углерода, охлажденный и очищенный в газопромывателе, используемый для сатурации сока, клеровки тростникового сахара-сырца.
Сатурация сахаросодержащего раствора – взаимодействие дефекованного сахаросодержащего раствора с диоксидом углерода сатурационного газа до заданного значения рН и щелочности, сопровождающееся образованием осадка карбоната кальция, с целью удаления несахаров путем адсорбции на осадке.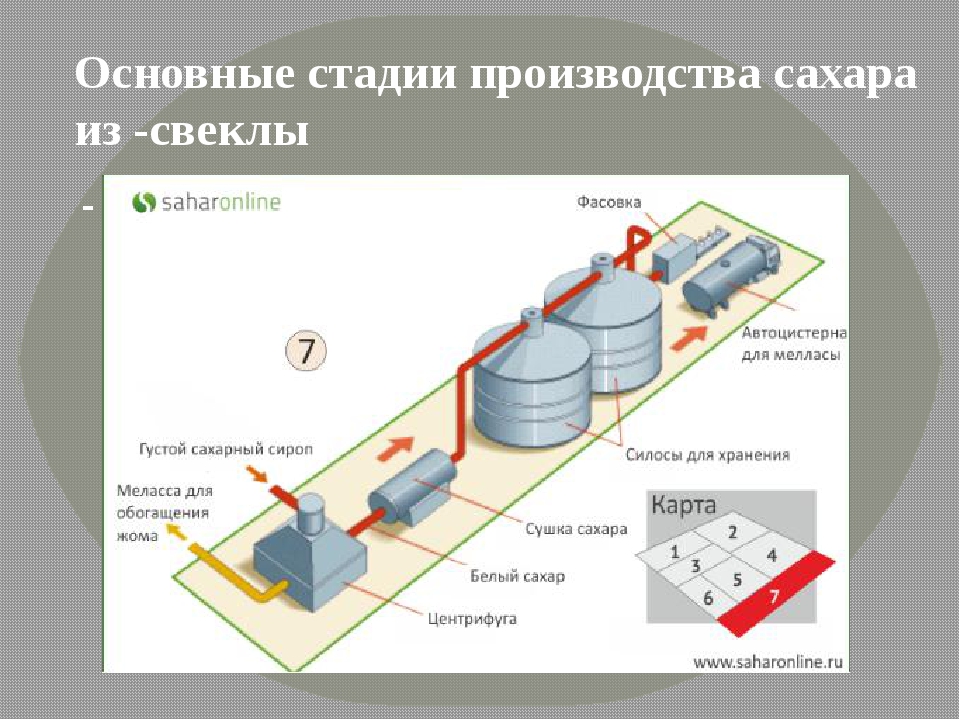
Сахар II кристаллизации или III кристаллизации – сахар, полученный в результате центрифугирования утфеля II кристаллизации или утфеля III кристаллизации.
Сахарный завод – промышленное предприятие, оснащенное технологической линией производства сахара с сопутствующей инфраструктурой.
Сахаристость сахарной свеклы – показатель, характеризующий массовую долю сахарозы в корнеплодах сахарной свеклы, выраженный в процентах к массе сахарной свеклы.
Сахарное производство – отрасль пищевой промышленности, промышленные предприятия которой производят сахар из первичного или вторичного сахароносного растительного сырья в результате выполнения комплекса последовательных технологических процессов и операций.
П р и м е ч а н и е – К первичному сахароносному сырью относится сахарная свекла, к вторичному – тростниковый сахар-сырец, сахар-песок и белый сахар.
Свеклосахарное производство – сахарное производство, в котором используется первичное сахароносное растительное сырье.
Свекловичная стружка – срезы сахарной свеклы определенных размеров и формы, получаемые путем изрезывания корнеплодов сахарной свеклы.
Сгущение очищенного сока – удаление воды из очищенного сока путем выпаривания до достижения заданного значения сухих веществ раствора.
Стандарт-сироп – концентрированный сахаросодержащий раствор заданной чистоты, используемый для уваривания маточного утфеля и утфеля I кристаллизации.
Сульфитация воды – взаимодействие воды с газообразным диоксидом серы, сернистой кислотой с целью достижения заданного значения рН.
Сульфитация сахарного раствора – взаимодействие сахарного раствора с газообразным диоксидом серы, сернистой кислотой или ее солями с целью предотвращения нарастания цветности и снижения вязкости при уваривании утфелей.
Сульфитация сока – взаимодействие сока с газообразным диоксидом серы, сернистой кислотой или ее солями с целью предотвращения нарастания цветности сока при выпаривании.
Сульфитированный сок – сок, обработанный газообразным диоксидом серы, сернистой кислотой или ее солями.
Сухие вещества продукта сахарного производства – твердые вещества, растворимые и нерастворимые в воде, содержащиеся в сырье и продуктах сахарного производства.
Сушка жома – удаление влаги из отжатого жома путем его высушивания до достижения нормируемого значения массовой доли влаги.
Сушка кристаллов сахара – совокупность процессов удаления влаги с поверхности кристаллов сахара и их охлаждения до достижения нормируемых значений массовой доли влаги и температуры.
Транспортерномоечная вода – смесь загрязненных вод после гидротранспортера и свекломойки.
Трехступенчатая кристаллизация сахарозы – совокупность технологических процессов и операций, объединяющих три последовательные стадии кристаллизации сахаросодержащего раствора с получением сахаров I, II и III кристаллизации.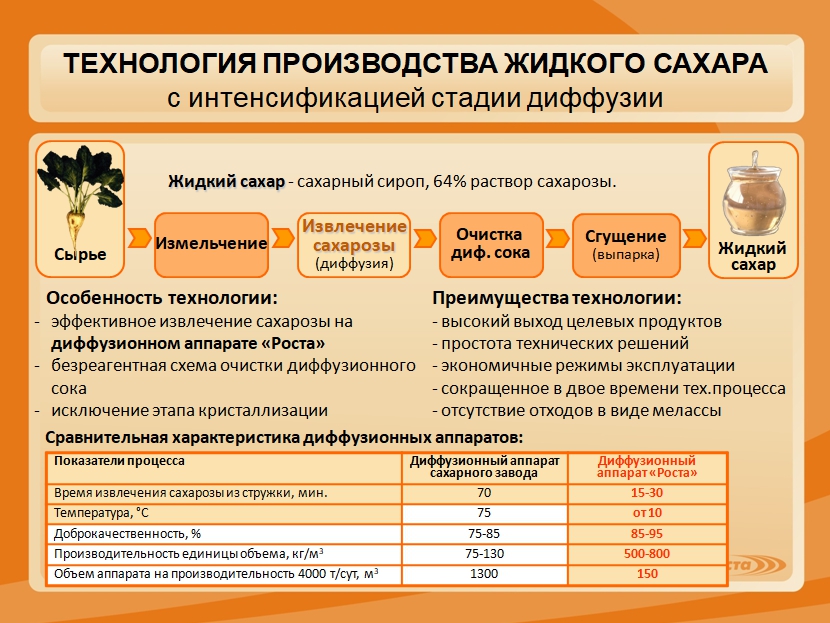
Уваривание утфеля – совокупность последовательных технологических процессов кристаллизации сахарозы, проводимых при кипении концентрированных сахаросодержащих растворов с образованием утфельной массы с заданным значением сухих веществ.
Утфель – масса, состоящая из кристаллов сахарозы и межкристального раствора.
Утфель I кристаллизации – утфель первой стадии последовательной кристаллизации сахарозы, предназначенный для получения белого сахара, сахара-песка.
Утфель II кристаллизации или III кристаллизации – утфель второй или третьей стадии последовательной кристаллизации, предназначенный для получения сахара II или III кристаллизации.
Хранение сахара – совокупность процессов, включающих складирование и поддержание на заданном уровне потребительских свойств сахара.
Хранение сахарной свеклы – совокупность операций, включающих складирование сахарной свеклы в кагаты и выполнение комплекса определенных технологических операций, направленных на сохранность исходных технологических свойств корнеплодов.
Цветность – показатель, характеризующий степень окрашенности продуктов сахарного производства, выраженный в условных единицах или единицах оптической плотности.
Центрифугирование утфеля – разделение утфеля на кристаллы сахарозы и межкристальный раствор под действием центробежных сил.
Чистота продукта сахарного производства – показатель, характеризующий отношение массовой доли сахарозы к сухим веществам продукта сахарного производства, выраженный в процентах.
Экстрагирование сахарозы – извлечение сахарозы из свекловичной стружки специально подготовленной водой с получением диффузионного сока.
Эффект кристаллизации – показатель, характеризующий степень выделения кристаллического сахара на стадии кристаллизации, определяемый как разность между чистотой продукта, поступающего на кристаллизацию, и чистотой межкристального раствора утфеля в конце стадии.
Сахар
Принципиальная схема производства сахара из свеклы представлена на рисунке ниже.
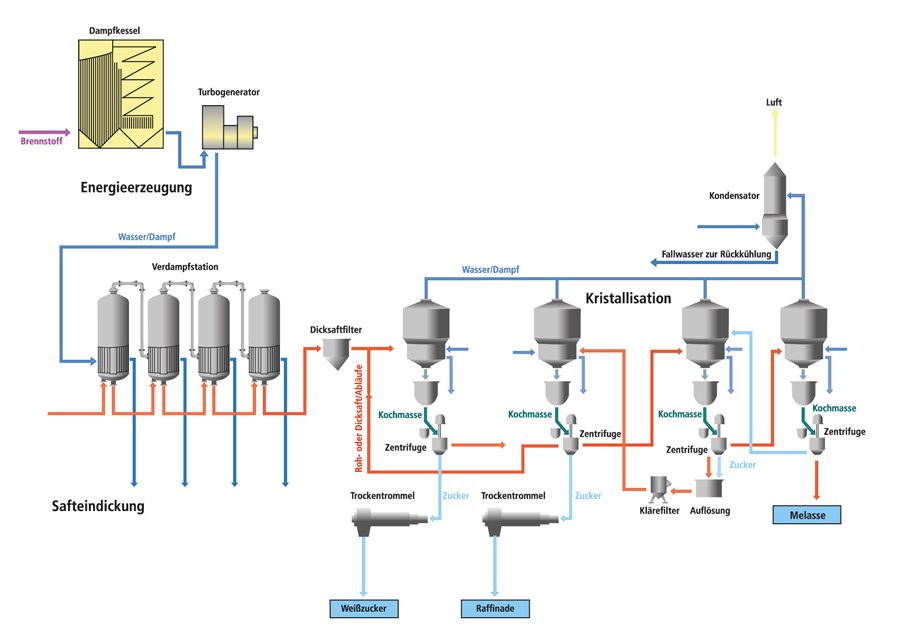
Сахарная свекла поступает на завод по гидравлическому транспортеру, на котором установлены ловушки для отделения легких и тяжелых примесей. С помощью свеклонасоса свекла подается в свекломойку, где она отмывается от земли. Отмытая свекла элеватором поднимается на высоту примерно 20 м к автоматическим весам, чтобы она дальше могла двигаться самотеком и тем самым сократить число транспортирующих механизмов. Взвешенная свекла затем для измельчения в стружку поступает на свеклорезки. Полученная стружка направляется в диффузионный аппарат, в котором происходит выделение сахара из стружки методом диффузии. Полученный в диффузионном аппарате диффузионный сок черного цвета, содержащий примерно 13% сахара и перешедшие в сок несахара, поступает на очистку.
Вначале его обрабатывают известью (известковым молоком). (Этот процесс получил название дефекации.) При такой обработке значительная часть несахаров, в первую очередь органических, выпадает в осадок. Дефекованный сок затем обрабатывают СО2 (сатурационным газом). Процесс обработки СО2 носит название сатурации. В процессе сатурации образуется карбонат кальция (СаСО3), который адсорбирует несахара и выпадает в виде осадка. Этот осадок отделяют фильтрованием. Фильтрат, так называемый сатурационный сок, содержит примерно 13% сахара и имеет светло-желтый цвет.
Процесс обработки СО2 носит название сатурации. В процессе сатурации образуется карбонат кальция (СаСО3), который адсорбирует несахара и выпадает в виде осадка. Этот осадок отделяют фильтрованием. Фильтрат, так называемый сатурационный сок, содержит примерно 13% сахара и имеет светло-желтый цвет.
Сатурационный сок обрабатывают SO2 (сульфитируют) с целью снижения цветности, а затем сгущают на выпарной установке до получения сиропа, содержащего примерно 60% сахара.
Из сиропа в вакуум-аппаратах проводят кристаллизацию сахара, получая утфель, представляющий собой смесь, состоящую примерно на 50% из кристаллов сахара и 50% сахарного раствора.
Утфель направляется на центрифуги, на которых из него выделяют кристаллический сахар и получают сахарный раствор (оттек). При пробеливании сахара водой получают два оттека. Кристаллический сахар сушат в сушилках, получая сахар-песок.
Для извлечения сахара из оттека его снова уваривают в вакуум-аппарате, получая утфель II. При разделении утфеля II на центрифугах получают желтый сахар I и оттек II. Оттек II уваривают и получают утфель III, при разделении которого получают желтый сахар II и мелассу.
При разделении утфеля II на центрифугах получают желтый сахар I и оттек II. Оттек II уваривают и получают утфель III, при разделении которого получают желтый сахар II и мелассу.
Желтые сахара растворяют (клеруют) и подвергают повторной кристаллизации в вакуум-аппаратах с целью получения сахара более высокого качества.
Меласса содержит примерно 50% сахара и дальнейшему обессахариванию не подвергается. Она используется в качестве сырья для производства спирта, дрожжей, лимонной, молочной и других пищевых кислот, а также в качестве корма для животных.
Побочными продуктами сахарного производства является обессахаренная стружка (жом) и фильтрационный осадок. В обессахаренной стружке содержится 0,3-0,5% сахара. Ее подвергают прессованию до содержания примерно 20% сухих веществ и используют в качестве корма для животных. Фильтрационный осадок используется в качестве удобрения.
версия для печати
Сладкий яд нашей жизни: история сахара | Культура и стиль жизни в Германии и Европе | DW
Сахарный тростник человек выращивает с древнейших времен. Об этом говорят находки археологов. Плантации сахарного тростника существовали уже 10 тысяч лет назад в Полинезии, 8 тысяч лет назад — в Индии. Так что не удивительно, что слово «сахар» пришло из древнеиндийского языка. А в VII веке н. э. в Персии научились получать из тростника сахар. Технология, в сущности, уже тогда была та же, что используется и сегодня: из стеблей тростника вываривали сахарный сироп, очищали его с помощью чаще всего гашеной извести или угля и разливали в деревянную или глиняную посуду. Жидкость испарялась, образовывались кристаллики сахара.
Об этом говорят находки археологов. Плантации сахарного тростника существовали уже 10 тысяч лет назад в Полинезии, 8 тысяч лет назад — в Индии. Так что не удивительно, что слово «сахар» пришло из древнеиндийского языка. А в VII веке н. э. в Персии научились получать из тростника сахар. Технология, в сущности, уже тогда была та же, что используется и сегодня: из стеблей тростника вываривали сахарный сироп, очищали его с помощью чаще всего гашеной извести или угля и разливали в деревянную или глиняную посуду. Жидкость испарялась, образовывались кристаллики сахара.
Богатые патриции Древнего Рима уже знали и ценили сахар, который ввозили из Индии через Египет. Для всех остальных жителей империи сахар заменял гораздо более дешевый и доступный сироп из прокипяченного виноградного сока. Люксовым товаром сахар был и в средневековой Европе, куда снова, после многовекового перерыва, попал благодаря крестоносцам. Сахар оставался предметом роскоши, и даже к королевскому столу его подавали только в особых случаях.
Как немцы научились получать сахар из сахарной свеклы
В России первая «сахарная палата» появилась в начале XVIII века при Петре Первом. Сахар был доступен немногим, и сырье для него оставалось в России еще в течение двухсот лет импортным товаром, когда его стали получать из отечественной сахарной свеклы.
Впервые эту технологию разработал и описал в 1747 году прусский химик, член Берлинской академии наук Андреас Сигизмунд Маргграф (Andreas Sigismund Marggraf), а промышленное производство сахара из корнеплода сахарной свеклы впервые стал налаживать под Берлином его ученик, выходец из семьи бежавших из Франции гугенотов Франц Карл Ашар (Franz Carl Achard). Интересно, что настоящее развитие это производство получило как раз благодаря французам. Прусский король просьбу Ашара о дополнительных инвестициях отклонил, а вот Наполеон деньги на это дал. Объявивший континентальную блокаду Англии император, среди прочего, всячески поощрял независимость от британского колониального тростникового сахара (к тому же очень дорогого). Наполеон даже раздавал земли крестьянам под выращивание сахарной свеклы.
Наполеон даже раздавал земли крестьянам под выращивание сахарной свеклы.
Сахарный тростник
Однако и по сегодняшний день сахарный тростник остается в мире главным сырьем для производства сахара. В 2016 году (это последние данные) в мире было произведено около 277 миллионов тонн сахарной свеклы и почти 2 миллиарда тонн сахарного тростника. Его выращивают, в основном, в Бразилии, Индии и Китае, тогда как ведущими производителями сахарной свеклы являются Россия, Франция и США.
Сахарная блокада США и кубинский сахар в СССР
В свое время американцы почти на 100 процентов импортировали свой сахар с Кубы. Фидель Кастро, решив наказать империалистов за антикубинскую политику, объявил США сахарную блокаду. Чем страшно обрадовал американских фермеров. Уже через год в стране выращивали свою сахарную свеклу, и построенные при поддержке государства заводы по ее переработке полностью обеспечивали США отечественным сахаром. А Кастро некуда стало экспортировать тростниковый сахар, — разве что продавать по дешевке на мировом рынке.
Помог Советский Союз. Люди старшего возраста помнят появившийся в начале 1960-х годов во всех магазинах и столовых светло-коричневый кубинский тростниковый сахар — странноватого вкуса, а главное — не сладкий. Народ даже ядовитую песенку сочинил на мотив известной тогда песни «Куба, любовь моя»:
«Куба, отдай наш хлеб,
Куба, возьми свой сахар…»
Сахарный яд
Сегодня в Европе, в среднем, потребляется примерно 37 кг сахара на человека в год. Разумеется, не только, так сказать, в чистом виде, но и в составе самых разных продуктов питания и напитков. В Германии на душу населения приходится 33,8 кг сахара в год. Надо сказать, что цифра эта весьма условная, так как не учитывает экспорта тех пищевых продуктов, которые производятся в Германии, но съедаются и выпиваются в других странах или годами, как вино, остаются в погребах или на полках магазинов. В них тоже есть сахар — причем, не только в тортах, конфетах, шоколадках, но также, например, в горьком пиве.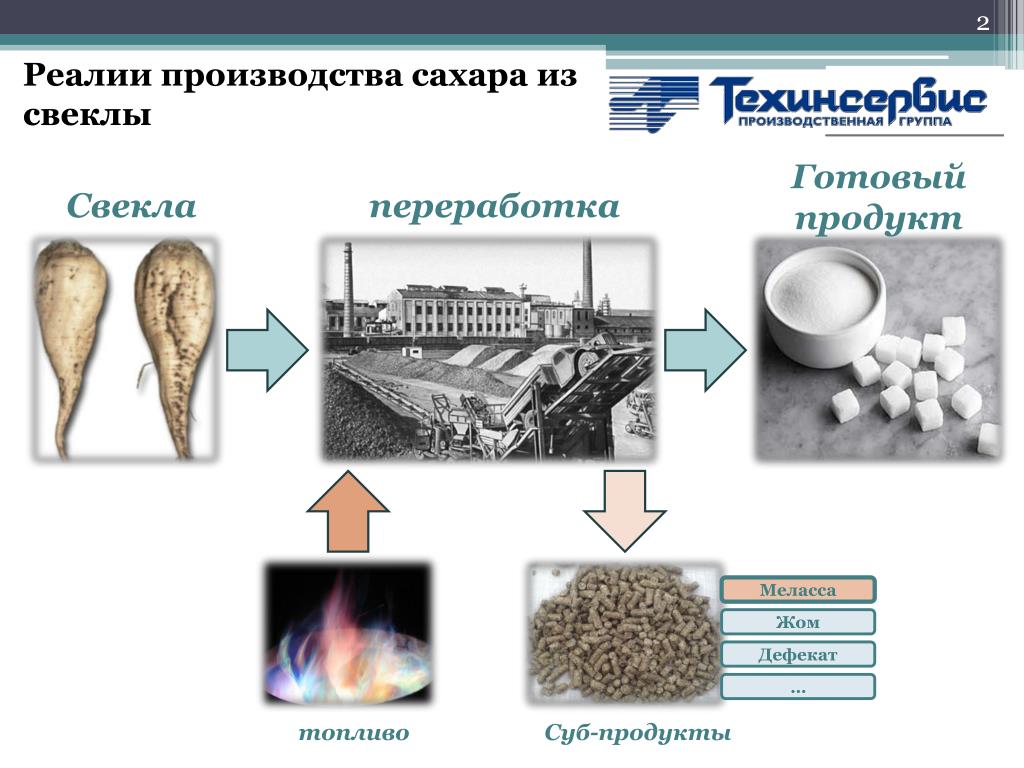 В общем, реальное потребление гораздо меньше.
В общем, реальное потребление гораздо меньше.
В России потребление сахара, по данным прошлого года, составляет почти 40 кг на человека. Это очень много: в полтора с лишним раза больше нормы, рекомендованной российским Минздравом. К предупреждениям медиков надо относиться серьезно. Давным-давно прошли те времена, когда сахар считался лекарством и продавался в аптеках. Сегодня к нему относятся намного более критично.
Уже в младенческом возрасте избыток сахара нередко способствует возникновению пищевой аллергии. Ну, а о риске заболевания сахарным диабетом, которому подвержены в значительной мере сладкоежки, сегодня знают все, — как и о зубном кариесе. У пожилых людей избыток сахара способствует быстрому развитию атеросклероза, вносит дезорганизацию в функции клеток. Не случайно сахар сегодня часто называют сладким ядом.
И все-таки: как же жить без него?..
Смотрите также:
Горькие факты о сахаре
Горькие факты о сахаре
Одно из самых опасных последствий злоупотребления сахаром и продуктами, его содержащими (например конфетами, шоколадом и пирожными), — диабет 2-го типа.
 Это эндокринное заболевание врачи обычно так и называют: сахарный диабет. Сегодня в мире почти 200 миллионов людей, страдающих диабетом. Больные с тяжелой формой нуждаются в постоянных инъкцих инсулина.
Это эндокринное заболевание врачи обычно так и называют: сахарный диабет. Сегодня в мире почти 200 миллионов людей, страдающих диабетом. Больные с тяжелой формой нуждаются в постоянных инъкцих инсулина.Горькие факты о сахаре
Нередко наш мозг реагирует на сахар примерно так же, как на алкоголь и другие вещества, способные вызывать зависимость. Попробуйте несколько дней полностью воздержаться от сладкого! Это, наверняка, будет непросто.
Горькие факты о сахаре
Одно из самых распространенных заблуждений, связанных с сахаром: коричневый сахар «здоровее» белого рафинада, более полезен для организма. Увы, это не так. Скажем, в 100 граммах и того, и другого вида сахара — 400 килокалорий, для зубов одинаково вредны и белый, и коричневый. Короче говоря, «оба хуже».
Горькие факты о сахаре
Сахар наш организм усваивает в два раза быстрее, а иногда и в пять раз быстрее, чем крахмал, превращая его в жиры.
Это значит, что сладостями мы буквально подкармливаем наши жировые клетки. Люди, которые едят много сладкого, нередко страдают от избыточного веса. Кроме того, в жиры наша печень перерабатывает и фруктозу, что может привести к сахарному диабету.Горькие факты о сахаре
У любителей сладкого кожа стареет быстрее. Чрезмерное употребление сахара вызывает реакцию организма, которую называют гликированием. Это «засахаривание» кожных тканей. Молекулы сахара приклеиваются к коллагеновым волокнам, которые теряют при этом естественную эластичность и способность выводить шлаки и яды из организма, что ведет к ускорению процесса старения клеток.
Горькие факты о сахаре
Многочисленные исследования немецких ученых подтверждают, что излишнее употребление сахара повышает и риск заболевания болезнью Альцгеймера. Высокие показатели содержания сахара в крови у пациентов с сахарным диабетом могут стать виновниками и нейродегенеративных заболеваний.
Горькие факты о сахаре
Любители сладкого склонны быстрее впадать в агрессию. Известно, что симптомы синдрома дефицита внимания и гиперактивности (СДВГ) у детей и подростков усиливаются от употребления сахара. Они становятся беспокойными, менее концентрированными и на любые мелочи реагируют более агрессивно.
Горькие факты о сахаре
Здоровая микрофлора кишечника защищает желудочно-кишечный тракт от влияния вредоносных бактерий и оказывает всяческую поддержку пищеварительному процессу. Чем больше сахара попадает в кишечник, тем легче непрошенные «гости» могут в нем размножаться. Грибки и паразиты обожают сахар! Особенно сandida albicans имеет обыкновение досаждать нам, провоцируя пресловутое «расстройство желудка».
Горькие факты о сахаре
Излишний сахар отягощает работу иммунной системы, отвечающей за борьбу с возбудителями различных заболеваний. Вскоре после излишней порции сладкого наш иммунитет функционирует на 40 процентов слабее, и мы рискуем «поймать» какое-нибудь воспаление.
Ведь сахар поглощает и столь необходимый нам витамин С.Автор: Инга Ваннер
 Это эндокринное заболевание врачи обычно так и называют: сахарный диабет. Сегодня в мире почти 200 миллионов людей, страдающих диабетом. Больные с тяжелой формой нуждаются в постоянных инъкцих инсулина.
Это эндокринное заболевание врачи обычно так и называют: сахарный диабет. Сегодня в мире почти 200 миллионов людей, страдающих диабетом. Больные с тяжелой формой нуждаются в постоянных инъкцих инсулина. Это значит, что сладостями мы буквально подкармливаем наши жировые клетки. Люди, которые едят много сладкого, нередко страдают от избыточного веса. Кроме того, в жиры наша печень перерабатывает и фруктозу, что может привести к сахарному диабету.
Это значит, что сладостями мы буквально подкармливаем наши жировые клетки. Люди, которые едят много сладкого, нередко страдают от избыточного веса. Кроме того, в жиры наша печень перерабатывает и фруктозу, что может привести к сахарному диабету.
 Ведь сахар поглощает и столь необходимый нам витамин С.
Ведь сахар поглощает и столь необходимый нам витамин С._________________________
Хотите читать нас регулярно? Подписывайтесь на наши VK-сообщества «DW на русском» и «DW Учеба и работа» и на Telegram-канал «Что там у немцев?»
Особенности производства и технология производства сахара из свеклы. Технология производства сахара.
Технология производства сахара из свеклы относится к непрерывно-поточному механизированному производству с высоким уровнем автоматизации основных процессов. Особенностью территориального размещения сахарных заводов является их жёсткая привязка к посевным площадям сахарной свеклы
Технология производства сахара из свеклы относится к непрерывно-поточному механизированному производству с высоким уровнем автоматизации основных процессов. Особенностью территориального размещения сахарных заводов является их жёсткая привязка к посевным площадям сахарной свеклы, поскольку перевозка свеклы на сколь-нибудь значительные расстояния экономически неэффективна. В ряде случаев, сахарные заводы имеют собственные посевные площади, расположенные непосредственно вблизи предприятия. Отходы сахарной промышленности (жом, барда, дефекационная грязь) могут быть использованы как удобрения, в некоторых случаях — и как корм для скота. Сахар – высококалорийная пища; его энергетическая ценность составляет около 400 ккал на 100г. Он легко переваривается и легко усваивается организмом, то есть служит концентрированным и быстро мобилизуемым источником энергии. С химической точки зрения сахаром можно назвать любое вещество из обширной группы водорастворимых углеводов. В быту же сахаром принято называть обычный пищевой подсластитель – сахарозу, сладкое кристаллическое вещество, выделяемое главным образом из сока сахарного тростника или сахарной свеклы. В чистом (рафинированном) виде сахар белый, а кристаллы его бесцветны.
В ряде случаев, сахарные заводы имеют собственные посевные площади, расположенные непосредственно вблизи предприятия. Отходы сахарной промышленности (жом, барда, дефекационная грязь) могут быть использованы как удобрения, в некоторых случаях — и как корм для скота. Сахар – высококалорийная пища; его энергетическая ценность составляет около 400 ккал на 100г. Он легко переваривается и легко усваивается организмом, то есть служит концентрированным и быстро мобилизуемым источником энергии. С химической точки зрения сахаром можно назвать любое вещество из обширной группы водорастворимых углеводов. В быту же сахаром принято называть обычный пищевой подсластитель – сахарозу, сладкое кристаллическое вещество, выделяемое главным образом из сока сахарного тростника или сахарной свеклы. В чистом (рафинированном) виде сахар белый, а кристаллы его бесцветны.
Буроватая окраска некоторых его сортов объясняется примесью мелассы – сгущенного растительного сока, обволакивающего кристаллы. Возникновение сахарной промышленности в России относится к началу 17 века, когда в Петербурге был построен первый сахарорафинадный завод, перерабатывающий привозной тростниковый сахар – сырец. Их сахарной свеклы сахар стали вырабатывать в России в начале 19 века. На сегодняшний день в России существует 96 сахарных заводов, работающих из которых только 84. На размещение предприятий сахарной промышленности решающее воздействие оказывает сырьевой фактор, т.е. сахарные заводы привязаны к посевным площадям сахарной свеклы, поскольку перевозка свеклы на сколько-нибудь значительные расстояния экономически неэффективна. Лидерами по производству сахара в нашей стране является Белгородская, Тамбовская, Воронежская и Липецкая области. Главный компонент сахара – сахароза. Это дисахарид, состоящий из глюкозы и фруктозы, не обладающие редуцирующими свойствами. В свекле содержится 25-28 % сухих веществ, из них на долю сахарозы приходится в среднем 17,5%. Сахароза растворена в соке, который заполняет вакуум клеток. Основным потребителем сахара в нашей стране остается население (около 55%), примерно 30% выпущенного отраслью продукции потребляется пищевой промышленностью.
Их сахарной свеклы сахар стали вырабатывать в России в начале 19 века. На сегодняшний день в России существует 96 сахарных заводов, работающих из которых только 84. На размещение предприятий сахарной промышленности решающее воздействие оказывает сырьевой фактор, т.е. сахарные заводы привязаны к посевным площадям сахарной свеклы, поскольку перевозка свеклы на сколько-нибудь значительные расстояния экономически неэффективна. Лидерами по производству сахара в нашей стране является Белгородская, Тамбовская, Воронежская и Липецкая области. Главный компонент сахара – сахароза. Это дисахарид, состоящий из глюкозы и фруктозы, не обладающие редуцирующими свойствами. В свекле содержится 25-28 % сухих веществ, из них на долю сахарозы приходится в среднем 17,5%. Сахароза растворена в соке, который заполняет вакуум клеток. Основным потребителем сахара в нашей стране остается население (около 55%), примерно 30% выпущенного отраслью продукции потребляется пищевой промышленностью.
Производство и потребление сахара носит сезонный характер. Свекловичный сахар в основном производится в сентябре – октябре после снятия урожая свеклы, сырцовый – в марте – июле. Пик потребления, как правило, приходится на июль, в пору массовых сельскохозяйственных заготовок. В зависимости от технологии производства сахар получается сыпучим или твердым (кусковым, колотым, леденцовым). САХАРНЫЙ ПЕСОК (сыпучий сахар) – его также называют «дробленым», «молотым», «гранулированным» или «сахарным песком» важен в кулинарии больше, чем любой другой: именно его чаще всего используют как подсластитель различных блюд. КУСКОВОЙ, КОЛОТЫЙ, ПИЛЕНЫЙ САХАР. «Кусковым» называют сахар, спрессованный в небольшие кубики. Рафинированный кусковой сахар называют «рафинадом». Кусковой сахар быстро растворяется в горячей воде, поэтому его очень часто подают к чаю. Чуть дольше растворяется в воде «колотый» или «пиленый» сахар – он, в сущности, представляет собой распиленный на маленькие части большой кусок сахара. ЛЕДЕНЦОВЫЙ, КАМЕННЫЙ САХАР внешне очень похожи на карамель (это полупрозрачное очень твердые кристаллы неправильной формы), да и процесс производства этого продукта очень напоминает приготовление «сосалок».
Свекловичный сахар в основном производится в сентябре – октябре после снятия урожая свеклы, сырцовый – в марте – июле. Пик потребления, как правило, приходится на июль, в пору массовых сельскохозяйственных заготовок. В зависимости от технологии производства сахар получается сыпучим или твердым (кусковым, колотым, леденцовым). САХАРНЫЙ ПЕСОК (сыпучий сахар) – его также называют «дробленым», «молотым», «гранулированным» или «сахарным песком» важен в кулинарии больше, чем любой другой: именно его чаще всего используют как подсластитель различных блюд. КУСКОВОЙ, КОЛОТЫЙ, ПИЛЕНЫЙ САХАР. «Кусковым» называют сахар, спрессованный в небольшие кубики. Рафинированный кусковой сахар называют «рафинадом». Кусковой сахар быстро растворяется в горячей воде, поэтому его очень часто подают к чаю. Чуть дольше растворяется в воде «колотый» или «пиленый» сахар – он, в сущности, представляет собой распиленный на маленькие части большой кусок сахара. ЛЕДЕНЦОВЫЙ, КАМЕННЫЙ САХАР внешне очень похожи на карамель (это полупрозрачное очень твердые кристаллы неправильной формы), да и процесс производства этого продукта очень напоминает приготовление «сосалок». Растворяется он гораздо дольше, чем кусковой.
Растворяется он гораздо дольше, чем кусковой.
Технология производства сахара
Сахарное производство нашей страны является крупной отраслью пищевой промышленности, объединяющей производство сахара-песка и сахара-рафинада.
Сырьем для производства сахара-песка служат или сахарная свекла, или сахарный тростник. Сахарный тростник принадлежит к семейству злаковых и возделывается на Кубе, в Мексике, Индии, Австралии и других странах с жарким климатом. Сахар, в основном сахароза, содержится в соке стеблей (12…15%) высотой до 4 м и толщиной до 50 мм. С 1 га сахарного тростника получается в два раза больше сахара, чем из свеклы.
Отжатый тростниковый сок очищают, уваривают и выделяют сахар-сырец. Тростниковый сахар-сырец – вещество светло-кремового цвета, массовая доля сахара в котором составляет 97…98%, инвертного сахара 0,6…0,8%, влаги 0,5…0,8%.
Сахарная свекла принадлежит к семейству маревых. Это двухлетнее засухоустойчивое растение. В первый год вырастает корнеплод, в во второй – стебель, цветы и семена. Для производства сахара используют корнеплоды первого года. Масса корнеплода 200…500 г. В корнеплоде массовая доля воды составляет 75%, сухих веществ, которые состоят из сахаров и несахаров. 25%.
Для производства сахара используют корнеплоды первого года. Масса корнеплода 200…500 г. В корнеплоде массовая доля воды составляет 75%, сухих веществ, которые состоят из сахаров и несахаров. 25%.
Содержание сахарозы в свекле, или сахаристость, может колебаться от 15 до 22% (в среднем 17,5%). Важным показателем свеклосахарного производства является доброкачественность или чистота сока, которая определяется отношением содержания сахарозы к содержанию сухих веществ, выраженным в %.
Период уборки сахарной свеклы составляет 40…50 сут. Сахарные заводы работают 110…150 сут в году, поэтому около 60% уб-ранной свеклы приходится закладывать на хранение. Хранение осуществляют в трапецеидальных кучах, называемых кагатами.
1. Доставка свеклы на завод и отделение примесей
Для обеспечения бесперебойной работы предприятия на нем создается 1…2-суточный запас свеклы, для чего предусматривают железобетонную емкость, так называемую бурачную, расположенную рядом с главным корпусом.
Из бурачной свекла подается на производство с помощью гидротранспортера – наклонного желоба, по которому свекла транспортируется водой. Подача воды составляет 600…700% от массы свеклы. В свою очередь. свекла содержит 5…15% примесей (ботвы, песка, камней, земли). Поэтому гидротранспортер оснащен песколовушками, ботволовушками и камнеловушками, которые улавливают как всплывшие на поверхность воды вещества (ботву, солому и др.), так и погружающиеся на дно (песок, камни и др.).
2. Мойка, взвешивание и изрезание свеклы
При подаче свеклы с помощью гидротранспортера часть механических примесей отделяется, но остаются примеси в виде прилипшей земли и др. Для их удаления свеклу подают в моечное отделение завода. Процесс мойки должен производиться очень тщательно, так как оставшиеся примеси ухудшают работу свеклорезок и загрязняют диффузионный сок. Для мойки свеклы применяют свекломоечные машины различных типов.
После моечных машин свеклу поднимают в верхнее отделение завода на высоту до 20 м, чтобы обеспечить ее гравитационный спуск на автоматические весы и в свеклорезки.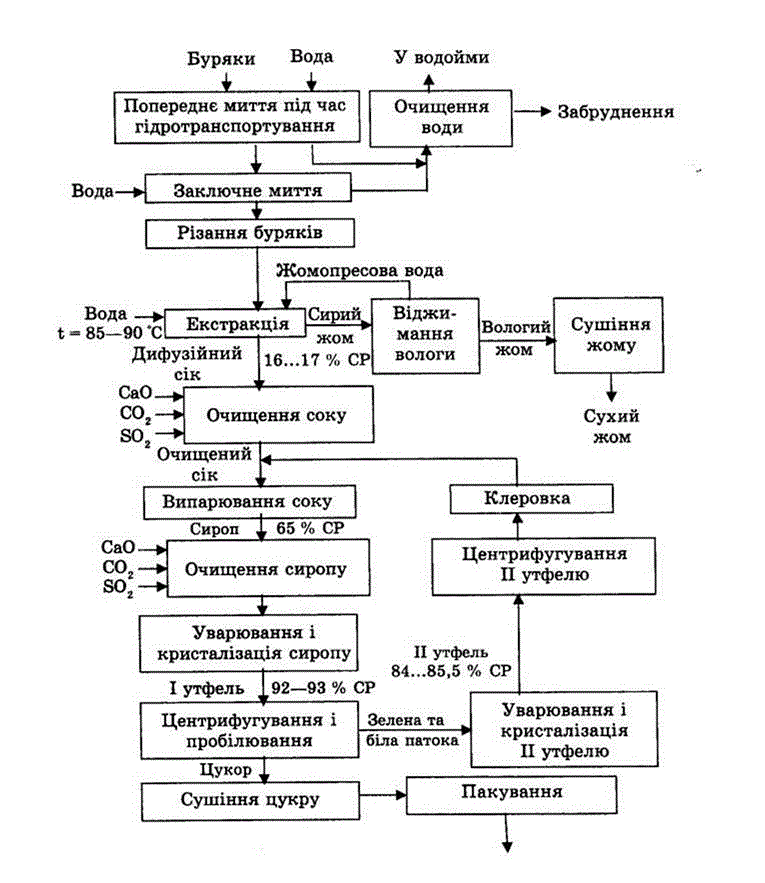 На транспортере свекла очищается от ферромагнитных примесей и подается в бункер автоматических весов для взвешивания.
На транспортере свекла очищается от ферромагнитных примесей и подается в бункер автоматических весов для взвешивания.
Сахар из свеклы извлекают диффузионным способом (растворением в воде). Для облегчения извлечения сахара свеклу измельчают в тонкую стружку желобчатой или пластинчатой формы. Толщина пластинок свекловичной стружки не должна превышать 0,5…1 мм, ширина полоски желобчатой стружки – 4…6 мм, пластинчатой – 2,5…3 мм.
Качество свекловичной стружки оказывает решающее влияние на работу диффузионного аппарата, служащего для извлечения сахара из стружки в водный раствор. Качество стружки оценивается длиной 100 г стружки, выраженной в метрах (число Силина), или отношением массы стружки длиной более 5 см к массе стружки менее 1 см (шведский фактор), для чего из определенной массы стружки выделяют частицы длиной менее 1 см и более 5 см. Для качественной стружки число Силина должно быть 9…15 м, а шведский фактор – не ниже 8.
Для изрезания свеклы наиболее распространены центробежные свеклорезки, в вырезах вертикальных цилиндрических корпусов которых неподвижно закреплено 12 или 16 ножевых рам.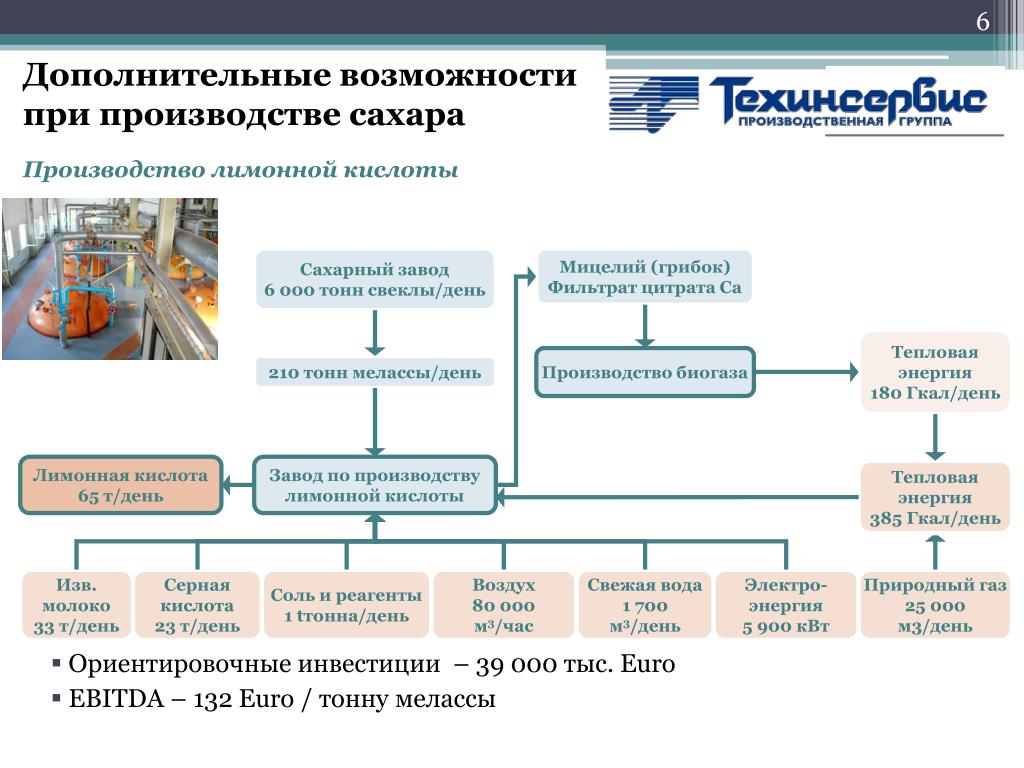 Свекла поступает во вращающийся ротор-улитку свеклорезки, вращается вместе с ротором, центробежной силой прижимается к ножам и режется. Затем свекловичная стружка по ленточному транспортеру поступает в отделение для получения диффузионного сока.
Свекла поступает во вращающийся ротор-улитку свеклорезки, вращается вместе с ротором, центробежной силой прижимается к ножам и режется. Затем свекловичная стружка по ленточному транспортеру поступает в отделение для получения диффузионного сока.
3. Получение диффузионного сока
Целью диффузионного процесса в сахарном производстве является извлечение из свекловичной стружки максимального количества сахарозы. Сахарозу из свеклы извлекают диффузионным способом (экстракцией), который заключается в противоточной обработке свекловичной стружки горячей водой. При этом сахароза и растворимые несахара переходят (диффундируют) в воду, в результате чего их содержание в стружке понижается, а в воде повышается. Такое движение растворимых веществ происходит под влиянием градиента концентрации. С повышением температуры диффузия ускоряется, поэтому процесс извлечения сахаров проводят при температуре 70…74°С. При более высокой температуре в раствор переходит часть пектиновых веществ. Таким образом, содержание сахаров в стружке в процессе экстракции снижается с 18,3% до 0,3%, а в диффузионном соке – повышается до 13,4%.
На отечественных сахарных заводах процесс извлечения сахарозы из свекловичной стружки осуществляется в непрерывнодействующих автоматизированных диффузионных установках. Длительность процесса диффузии составляет не более 80 мин, так как ее увеличение приводит к повышению содержания растворимых пектиновых веществ в диффузионном соке и его вязкости, а также к ухудшению условий дальнейшей очистки.
При понижении температуры ниже 70°С интенсивно развиваются микроорганизмы. С увеличением расхода воды на обессахаривание стружки снижаются потери сахаров в жоме, но на практике ограничивают его величиной 120…130% от массы стружки, идущей на диффузию, что обусловлено необходимостью экономии топлива, расходуемого на выпаривание лишней воды при сгущении сока.
Диффузионный сок является благоприятной средой для развития микроорганизмов, поступающих вместе со свеклой и водой. Развитие микроорганизмов подавляют улучшением отмывания свеклы, обеспечением чистоты диффузионной установки и питающей воды, а также ритмичной работой. Кроме того, в диффузионный аппарат периодически подают раствор формалина.
Кроме того, в диффузионный аппарат периодически подают раствор формалина.
В диффузионном соке, выходящем из диффузионного аппарата, содержится много мезги (мельчайших частиц стружки), ухудшающей дальнейшую переработку сока. Поэтому диффузионный сок перед подачей на дальнейшую переработку очищают от мезги.
Обессахаренная стружка (жом) выводится из верхней части диффузионного аппарата и подается в жомовый пресс. Массовая доля сухих веществ в стружке перед прессованием составляет примерно 8%. Жом после прессования в шнековых прессах имеет массовую долю сухих веществ 12…14%, если он в сыром виде будет скармливаться скоту, либо его прессуют до 22…25% сухих веществ и направляют на высушивание до массовой доли сухих веществ 86%. Для сохранения и повышения кормовой ценности жом обогащают добавками и брикетируют. В среднем выход сушеного жома составляет 4,5…5% от массы свеклы.
4. Очистка диффузионного сока
Из свеклы в диффузионный сок переходит почти вся сахароза и до 90% растворимых несахаров. Кроме того, в диффузионном соке содержится много мелких частиц свеклы (мезги), которые на воздухе быстро темнеют и пенятся.
Кроме того, в диффузионном соке содержится много мелких частиц свеклы (мезги), которые на воздухе быстро темнеют и пенятся.
Из такого сока без очистки трудно выделить сахарозу, так как несахара существенно замедляют скорость кристаллизации и увеличивают содержание сахара в отходах (мелассе). Одна часть несахаров удерживает в мелассе до 1,5 частей сахарозы. Чтобы получить максимальный выход сахара-песка и низкий выход мелассы, из диффузионного сока необходимо удалить как можно больше несахаров и довести его до слабощелочной реакции, в которой сахароза наиболее устойчива к разложению.
Производится довольно сложная и многоступенчатая очистка диффузионного сока.
Первой ступенью очистки диффузионного сока является преддефекация. При этом в диффузионный сок добавляют известковое молоко в количестве 0,2…0,3% СаО от массы свеклы равномерно во времени в течение 20…30 мин при температуре 40…60°С. Целью преддефекации является коагуляция (укрупнение) частиц коллоидной дисперсии для выведения их из раствора.
Далее производится основная дефекация. Целью основной дефекации является вторичная обработка диффузионного сока избытком извести сразу же после преддефекации. Основной дефекацией достигаются полное разложение амидов кислот, редуцирующих и пектиновых веществ, солей аммония, омыление жиров, а также создание избытка извести, необходимой для получения достаточного количества карбоната кальция при дальнейшей очистке – на 1-й сатурации. Общее количество активной извести, расходуемой на преддефекацию и основную дефекацию, составляет 2,2…2,5% СаО от массы свеклы. Температура, длительность процесса и доза известкового молока определяются лабораторией в зависимости от качества перерабатываемой в данный момент свеклы.
Сразу же после дефекации производится 1-я сатурация. После основной дефекации нефильтрованный сок, содержащий известь (меньшую часть – в растворе и большую часть – в осадке), поступает на 1-ю сатурацию, где его обрабатывают сатурационным газом (смесью газов, содержащих в большом количестве диоксид углерода). Диоксид углерода (СО2) вступает в реакцию с гидроксидом кальция (Са(ОН)2) и образует карбонат кальция (СаСО3). На положительно заряженной поверхности свежеобразованных кристаллов СаСО3 адсорбируются несахара сока, в том числе продукты распада пектиновых и других веществ, несущих отрицательный заряд.
Диоксид углерода (СО2) вступает в реакцию с гидроксидом кальция (Са(ОН)2) и образует карбонат кальция (СаСО3). На положительно заряженной поверхности свежеобразованных кристаллов СаСО3 адсорбируются несахара сока, в том числе продукты распада пектиновых и других веществ, несущих отрицательный заряд.
Таким образом, если на предварительной и основной дефекациях химическая очистка осуществлялась путем коагуляции, осаждения и разложения несахаров, то на 1-й сатурации проводится физико-химическая очистка сока адсорбцией, что и является основной целью 1-й сатурации. Кроме того, образующийся кристаллический осадок СаСО3 служит основой для создания фильтрующего слоя при фильтрации сока.
Образовавшийся осадок СаСО3 с адсорбированными несахарами отфильтровывают.
Нефильтрованный сок 1-й сатурации содержит 4…5% взвешенных частиц, которые необходимо отделить, чтобы продолжить дальнейшую очистку сока. Фильтрование чаще всего производят на листовых саморазгружающихся фильтрах-сгустителях периодического действия..files/image031.gif) Сок 1-й сатурации на фильтре разделяется на фильтрованный сок и сгущенную суспензию.
Сок 1-й сатурации на фильтре разделяется на фильтрованный сок и сгущенную суспензию.
Отфильтрованный сок содержит на выходе из фильтра не более 1 г/л твердой фазы и без контрольного фильтрования направляется на 2-ю сатурацию. Сгущенную суспензию сока 1-й сатурации повторно фильтруют в камерных вакуум-фильтрах, на которых осадок промывается горячей водой и просушивается воздухом или паром. Разбавленный сок, получаемый на первой стадии промывки, присоединяют к отфильтрованному соку. Сильно разбавленный сок, получаемый на окончательной стадии промывки осадка, используют в других технологических процессах. В фильтрованном осадке содержится 75…80% карбоната кальция и 20…25% органических и неорганических несахаров. Он используется в сельском хозяйстве для известкования кислых почв. Потери сахарозы в фильтрованном осадке составляют примерно 1% от его массы. На промывку фильтрованного осадка расходуется 105…110% воды от массы осадка.
Тщательно отфильтрованный, чистый сок подвергают ^ 2-й сатурации для того, чтобы перевести оставшиеся после 1-й сатурации в растворе гидрооксиды кальция, калия и натрия в углекислые соли и вывести их в осадок, а также вывести в осадок кальций, связанный с органическими кислотами в комплексы.
Для повышения качества к соку перед 2-й сатурацией добавляют небольшое количество извести (0…0,5% СаО от массы сока), что способствует не только дополнительному разложению несахаров, но и увеличению адсорбционной поверхности в результате большего образования карбоната кальция. Перед 2-й сатурацией сок нагревают до температуры 93…95°С и в течение 10 мин сатурируют (продувают СО2). При сатурировании из сока испаряется более 1% воды, и он охлаждается на 2…5°С. Для дополнительного удаления кальция из раствора сок после сатуратора следует подвергнуть “дозреванию” в течение 10…15 мин при интенсивном перемешивании в отдельном сосуде, что снижает накипеобразование в выпарной установке.
После “дозревания” сок 2-й сатурации фильтруют на листовых фильтрах таким же образом, как и сок 1-й сатурации. Фильтрат направляют на сульфитирование, а сгущенную суспензию – на преддефекацию.
Сульфитацией называется обработка сахарных растворов диоксидом серы (SO2), который получают сжиганием комовой серы на воздухе в специальной печи. В получаемом сульфитационном газе содержится 10…15% SО2, воздуха 85…90%.
В получаемом сульфитационном газе содержится 10…15% SО2, воздуха 85…90%.
Диоксид серы – бесцветный газ с резким запахом, ядовит, вызывает удушье, хорошо растворим в воде, но только небольшая часть растворенного диоксида серы реагирует с водой, образуя сернистую кислоту.
Целями сульфитации являются: обесцвечивание сока, снижение его вязкости, а также обеззараживание. При пропускании сульфида серы через сок образуется сернистая кислота, являющаяся сильным восстановителем. Она частично переходит в серную кислоту; при этом выделяется молекулярный водород, который восстанавливает органические окрашенные вещества. Действие сернистого газа продолжается и при выпаривании, что способствует меньшему потемнению сиропа. Коэффициент использования диоксида серы составляет 70…80%, оптимальное значение рН сульфитированного сока составляет 8,5…8,8.
Повышение урожайности сахарной свеклы и улучшение ее технологического качества
Повышение урожайности сахарной свеклы и улучшение ее технологического качества — важнейшая задача свеклосахарного комплекса.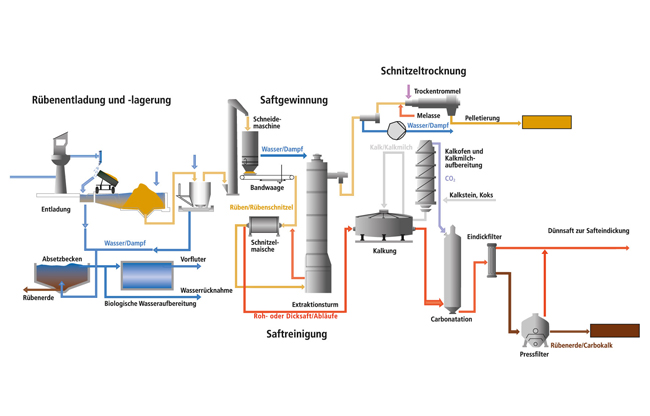
Решение этой задачи зависит от многих факторов: сорта возделываемой свеклы, качества используемых семян, качества почвы, применяемых агротехнических мероприятий и т. д.
Качество семян является одним из решающих факторов. Поэтому фирмы, занимающиеся производством семян, уделяют их качеству особое внимание, проводя соответствующую обработку семян и осуществляя тщательный их контроль. При этом, наряду с традиционными показателями качества семян (засоренности, всхожести и т. д.), в настоящее время определяются с помощью современного оборудования биологические, физиологические и другие показатели.
Кроме качества семян на урожайность большое влияние оказывает технология возделывания свеклы. В этом направлении в последние годы ведутся интенсивные разработки.
Так, в начале двадцатых годов на Украине В.С. Глуховским с сотрудниками разработан новый, так называемый гребневый способ возделывания сахарной свеклы. Способ заключается в том, что с осени готовят гребни, что способствует в этот период интенсивному накоплению влаги, а весной — ускоренному созреванию почвы в их зоне.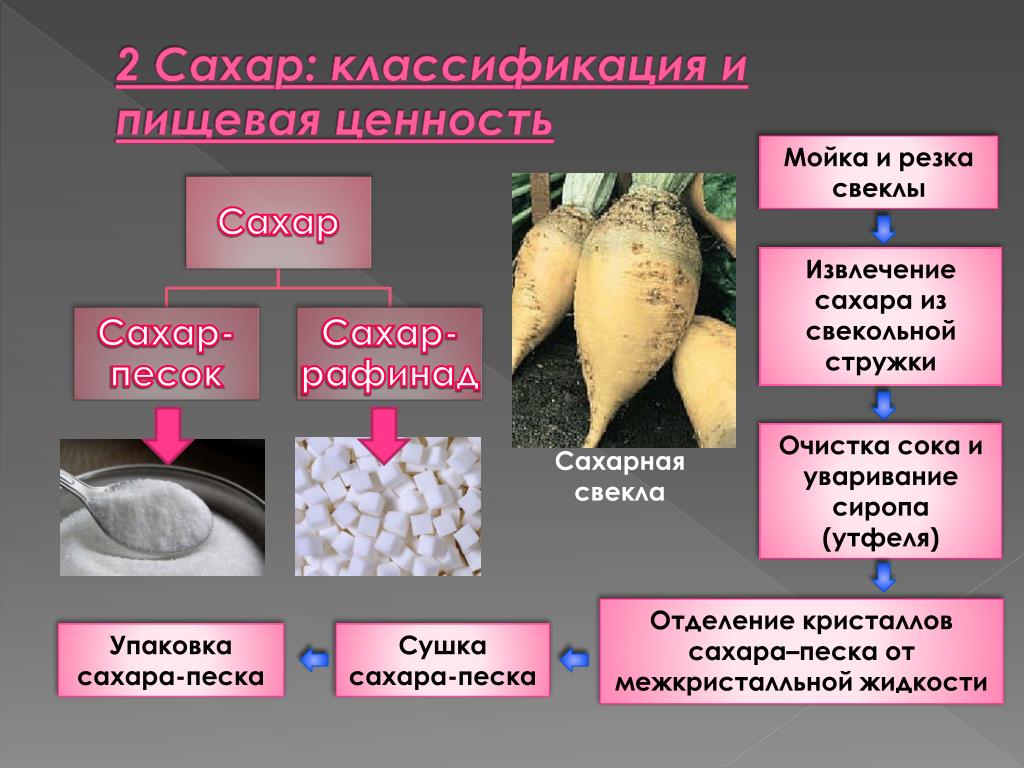 В гребни затем высевают семена. (Данная технология подобна технологии при выращивании картофеля.)
В гребни затем высевают семена. (Данная технология подобна технологии при выращивании картофеля.)
Преимуществом такой технологии является то, что вследствие большей поверхности гребня почва весной прогревается гораздо быстрее, что способствует более быстрому росту растений.
На грядках из-за большей рыхлости почвы свекла образует более длинные корешки и тем самым достигнет быстрее слоя почвы, содержащего влагу. На гребнях меньше потери влаги испарением из нижнего слоя. Гребневый способ возделывания свеклы способствует ускорению прорастания семян и росту растений.
Поскольку почва на гребнях менее уплотнена, то свекла образует меньше боковых корешков, ее легче убирать.
Гребневый способ повышает продуктивность сахарной свеклы. Об этом свидетельствует опыт применения этого способа в последние годы в Германии и Италии.
Так, на севере Германии в течение четырех лет при применении данного способа урожайность была примерно на 15% выше. В Италии, которая только начала применять этот способ, урожайность была на 30. ..45% выше.
..45% выше.
Недостаточно широкое распространение данной технологии возделывания свеклы может быть связано с отсутствием необходимого оборудования.
В начальный период для формирования грядок использовалась техника, которая применялась при возделывании картофеля. Однако она не всегда обеспечивала получение устойчивых грядок.
Фирмой Delitzsch в настоящее время разработана техника, позволяющая одновременно формировать 12 грядок и высевать в них семена. Это открывает возможности для более широкой реализации данного способа.
Уборка свеклы. Начало копки свеклы зависит от сроков пуска завода, убрать ее необходимо до наступления морозов. Уборка свеклы проводится поточным (с челночным) способом вывозки корнеплодов на завод или поточно-перевалочным способом. Для снижения потерь массы и сахара при необходимости хранения (более 2…3 суток) корнеплодов на перевалочных площадках в буртах их следует укрывать ботвой или соломой.
в промышленных и домашних условиях.

Привычная нам свекла имеет разновидность, переработка которой ведется в промышленных масштабах. Сегодня Россия занимает лидирующую позицию по производству из свеклы сахара. Как происходит производство продукта из сахарной свеклы и можно ли его приготовить дома?
От тростника к свекле
История получения сахара из растений уходит вглубь веков. Долгое время его производили из сахарного тростника. В середине 18 века ученый А. Маргграф в ходе исследования обнаружил, что такой же сахар содержится в свекле. Ответ на вопрос, сколько в свекле сахара, менялся. В то время его содержание в сахарной свекле не превышало 1,3%. Но селекционеры вывели сорта с 20-типроцентным количеством сахара в сахарной свекле.
История переработки сахарной свеклы в России началась в 19 веке и продолжается по сей день.
Промышленное производство сахара
Переработка сахарной свеклы и технология производства сахара на заводе – сложный и полностью автоматизированный процесс. Начинается он вне стен завода, когда выращенный овощ собирают и грузят в машины. Грузовики привозят сырье, после взвешивания выгружают в специальный отсек. Отсюда свекла по конвейерной ленте поступит в недра завода, выйдет из которого в виде нескольких продуктов: сахара, патоки, жмыха и удобрения.
Грузовики привозят сырье, после взвешивания выгружают в специальный отсек. Отсюда свекла по конвейерной ленте поступит в недра завода, выйдет из которого в виде нескольких продуктов: сахара, патоки, жмыха и удобрения.
Следующий этап – очистка корнеплодов от земли и ботвы. Они долгое время моются в специальных мойках, после чего поступают в цех для разрезания. Здесь агрегат превращает свеклу в ровную стружку, после чего она продолжает путешествие – перемещается в диффузионный аппарат. Это важный этап переработки, когда в процессе промывания водой свекольная стружка насыщает воду сахаром. Отработанный жмых постепенно выходит из переработки и отправляется на корм скоту. А сок продолжает переработку.
Процесс усложняется. Задача – удалить из сока примеси, которые мешают кристаллизации сахара. В сок добавляют известковое молоко, которое после нагревания переводит в осадок нежелательные примеси.
Далее очищенный сок выпаривают – так получается сахарный сироп. Он фильтруется и сгущается в специальных аппаратах.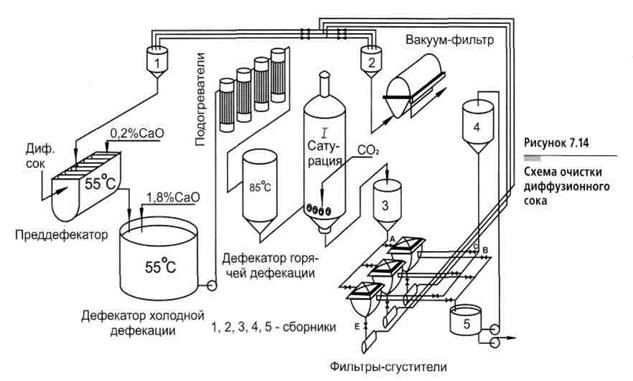 Но и это еще не все. Полученный сахарный песок отделяют от патоки в центрифугах. А чтобы он стал белым – его промывают и сушат. Последний этап – расфасовка. В процессе сложной переработки из одной тонны корнеплодов выход сахара составляет 100-150 кг.
Но и это еще не все. Полученный сахарный песок отделяют от патоки в центрифугах. А чтобы он стал белым – его промывают и сушат. Последний этап – расфасовка. В процессе сложной переработки из одной тонны корнеплодов выход сахара составляет 100-150 кг.
При покупке в магазине сахара обратите внимание, из чего, кем и когда он изготовлен. Вся информация должна быть на этикетке. От покупки стоит отказаться, если в производстве использовались генетически модифицированная свекла. Покупателей может насторожить диоксид серы, но без него не обходится любое производство сахара.
Производство в домашних условиях
Можно самостоятельно получить натуральный продукт без вредных примесей. Рассмотрим, как делают сахарный сироп дома.
Мытые корнеплоды варим в течение часа в эмалированной посуде, поддерживая кипящий режим. После остывания очищаем от кожуры и нарезаем тонкой соломкой. Нарезанные корнеплоды оборачиваем марлей и помещаем под пресс, чтобы отжался сок. Достаем из-под пресса жмых и заливаем горячей водой в соотношение один к двум. Даем полчаса отстояться, процеживаем жидкость и добавляем в нее ранее выжатый сок. Нагреваем его до 70-80 градусов, после чего начинаем выпаривать. В результате выпаривания должен получиться сахарный сироп.
Достаем из-под пресса жмых и заливаем горячей водой в соотношение один к двум. Даем полчаса отстояться, процеживаем жидкость и добавляем в нее ранее выжатый сок. Нагреваем его до 70-80 градусов, после чего начинаем выпаривать. В результате выпаривания должен получиться сахарный сироп.
Можно получить густой свекольный сироп другим способом, но для этого нужен автоклав или котел с решеткой у дна. Сначала так же отвариваем свеклу и очищаем от кожуры. Далее для распаривания помещаем на час в автоклав. Мягкие корнеплоды размельчаем и 2 раза прессуем. Сок, полученный в процессе 2-ух отжимов, выпариваем.
Свекольный сироп можно использовать для приготовления варенья и выпечки. Хранить его лучше в закатанных банках в темном и прохладном месте.
Приготовив дома свекольный сироп, вы будете уверены в его составе.
Все новости
ООО «Ромодановосахар» — по производству сахара самое большое предприятие в Приволжском федеральном округе и в десятке крупнейших заводов России. Это единственное предприятие в Республике Мордовия, которое производит свекловичный сахар. Производство продукции началось ещё в прошлом веке, в 1961 году. Этой осенью завод отмечает 60-летний юбилей. Возглавляет его вот уже 16 лет Александр Иванович Атласов.
Это единственное предприятие в Республике Мордовия, которое производит свекловичный сахар. Производство продукции началось ещё в прошлом веке, в 1961 году. Этой осенью завод отмечает 60-летний юбилей. Возглавляет его вот уже 16 лет Александр Иванович Атласов.
Новые технологии облегчают дело
Предприятие по производству сахара начинается со свёклоприёмного пункта. Сюда привозят сахарную свёклу все хозяйства республики, на чьих полях её выращивают.
— Надо сказать, на приёмном пункте весь процесс автоматизирован. Во время эпидемии коронавируса это очень важно, — говорит В.В. Захаркин, заместитель директора ООО «Ромодановосахар». Каждой машине, которая возит свёклу, выдают электронную пластиковую карточку — накладную, где вся информация о машине: номер, перевозчик (организация или ИП), имеется ли прицеп (самосвал разгружает назад или на бок, или с бортом). Это нужно для того, чтобы автоматика сама контролировала за машиной и посылала на разгрузку в нужное место.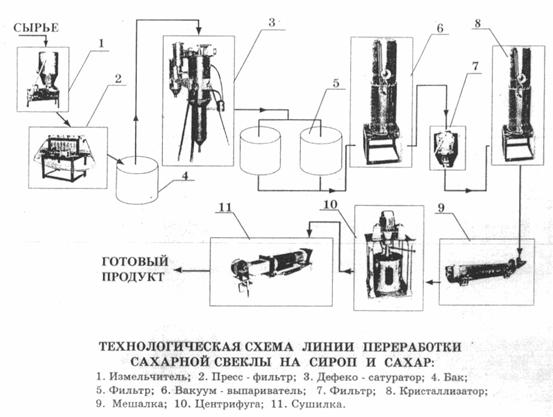 Когда машина загружается в поле свёклой, и там есть терминал, где каждому водителю на карточку отмечают всё необходимое: с какого поля взята продукция, номер автомашины, где находится, в каком хозяйстве. И когда он приезжает на завод, взвешивает вес гружёной машины и отправляется на нужный буртоукладчик, который выгружает сахарную свёклу. У механизатора буртоукладчика так же есть терминал, который отмечает — машина разгружена. Затем пустая машина едет вновь на весы, отмечается вес пустой машины, потом автоматически водителю выдают товарно-транспортную накладную. Кроме этого, как отметил В.В. Захаркин, если нужно машину направляют в лабораторию. Здесь лаборанты смотрят или каждую третью машину, или каждую десятую. Как рассказала работница сырьевой лаборатории — Betalauser системный оператор Надежда Евгеньевна Маньченкова, на территории лаборатории есть пробоотборник, который из кузова машины берёт свёклу и отдаёт в лабораторию, где определяют загрязнённость и сахаристость.
Когда машина загружается в поле свёклой, и там есть терминал, где каждому водителю на карточку отмечают всё необходимое: с какого поля взята продукция, номер автомашины, где находится, в каком хозяйстве. И когда он приезжает на завод, взвешивает вес гружёной машины и отправляется на нужный буртоукладчик, который выгружает сахарную свёклу. У механизатора буртоукладчика так же есть терминал, который отмечает — машина разгружена. Затем пустая машина едет вновь на весы, отмечается вес пустой машины, потом автоматически водителю выдают товарно-транспортную накладную. Кроме этого, как отметил В.В. Захаркин, если нужно машину направляют в лабораторию. Здесь лаборанты смотрят или каждую третью машину, или каждую десятую. Как рассказала работница сырьевой лаборатории — Betalauser системный оператор Надежда Евгеньевна Маньченкова, на территории лаборатории есть пробоотборник, который из кузова машины берёт свёклу и отдаёт в лабораторию, где определяют загрязнённость и сахаристость.
— Лаборанты моют свёклу, очищают и она поступает ко мне, — говорит Н. Е. Маньченкова. Я уже определяю сахаристость. Если этот показатель максимально высокий, т.е. очень хорошие технологические показатели свёклы, то и выход сахара будет больше.
Е. Маньченкова. Я уже определяю сахаристость. Если этот показатель максимально высокий, т.е. очень хорошие технологические показатели свёклы, то и выход сахара будет больше.
Когда мы были на свёклоприёмном пункте, обратили внимание как много здесь продукции. Виктор Владимирович объяснил так:
— Из привезённой на приёмный пункт свёклы делаются производственные кагаты, проще говоря, бурты из корнеплодов. Здесь свёкла будет лежать несколько дней. Мы не знаем, какая будет погода. Могут пойти дожди, а работу завода не остановишь. Поэтому мы храним свёклу в буртах на территории предприятия. Если говорить откровенно, 60-70% сладких корнеплодов с поля напрямую поступает на производство. Это для нас очень хорошо. Ведь чем меньше потери сахаристости и веса, тем лучше. Потому что, сахарная свёкла, которая хранится на полях хозяйств, на много лучше сохраняет сахар, чем та, которая долго хранится на территории завода.
Выращивание сахарной свёклы
Производство завода начинается ещё с поля. Вырастить сахарную свёклу дело нелёгкое. Те, кто этим занимается, хорошо понимают, в настоящее время для этого нужны современная техника, технологии и люди, которые знают весь производственный процесс. В ООО «Ромодановосахар» это хорошо понимают, и поддерживают тесную связь с сельскохозяйственными предприятиями республики.
Вырастить сахарную свёклу дело нелёгкое. Те, кто этим занимается, хорошо понимают, в настоящее время для этого нужны современная техника, технологии и люди, которые знают весь производственный процесс. В ООО «Ромодановосахар» это хорошо понимают, и поддерживают тесную связь с сельскохозяйственными предприятиями республики.
— Чтобы сахарная свёкла давала хороший урожай, поля для посева надо начинать готовить за два года, — рассказывает В.В. Захаркин. Свёклу хорошо сеять после озимой пшеницы или по парам. Тогда на полях будет меньше сорняков. По потребности нужно использовать гербициды и вносить удобрения. Кроме этого, землю нужно хорошо пахать, культивировать, чтобы поле было ровным как стол. Это нужно для того, чтобы после посева свёкла взошла одновременно и выросла одинаковая. Если поле не ровное, свёкла вырастет в одном месте большая, в другом маленькая и вовремя уборки урожая корнеплоды порежутся. Урожайность будет ниже.
Чтобы вырастить хороший урожай сахарной свёклы надо подготовить для посева и хорошие семена. Как отметил В.В. Захаркин, завод вместе с одним из хозяйств в 2007 году создал сортоиспытательный участок, где высеваются и выращиваются 60, иногда 70 — 80 гибридов этой культуры как российской, так и зарубежной селекции. Мы выбираем те сорта-гибриды, которые подходят для нашего климата. На рынке около 300 гибридов, нам надо же выбрать десяток. Выбрать такие, которые дают большой урожай и высокую сахаристость.
Как отметил В.В. Захаркин, завод вместе с одним из хозяйств в 2007 году создал сортоиспытательный участок, где высеваются и выращиваются 60, иногда 70 — 80 гибридов этой культуры как российской, так и зарубежной селекции. Мы выбираем те сорта-гибриды, которые подходят для нашего климата. На рынке около 300 гибридов, нам надо же выбрать десяток. Выбрать такие, которые дают большой урожай и высокую сахаристость.
Как пояснил Виктор Владимирович в этом году сахарной свёклы посеяно 22 300 гектар, урожайность ожидается 280-300 центнеров с гектара и убрать её надо до 25-26 октября.
— Если убрать урожай вовремя это очень хорошо, — говорит Виктор Владимирович, в это время сахарная свёкла уже созрела, набрала нужную сахаристость, и готова к переработке.
Производство сахара — дело не лёгкое
Сладкую свёклу привозят на приёмный пункт, где сырьевая лаборатория проверяет её на качество. Так начинается не лёгкий технологический процесс производства сахарного песка. На завод свёкла поступает по гидравлическому транспортёру, где она промывается от грязи. Чистую свёклу автоматически поднимают на весы. Затем взвешенная свёкла поступает на резку. Резанная свёкла поступает в диффузионные аппараты, где идет процесс извлечения сахара. Из диффузных аппаратов выходит сравнительно тёмный сок, его пропускают через известковое молоко.
На завод свёкла поступает по гидравлическому транспортёру, где она промывается от грязи. Чистую свёклу автоматически поднимают на весы. Затем взвешенная свёкла поступает на резку. Резанная свёкла поступает в диффузионные аппараты, где идет процесс извлечения сахара. Из диффузных аппаратов выходит сравнительно тёмный сок, его пропускают через известковое молоко.
Как отметила главный технолог предприятия Валентина Вячеславовна Бычкова, после осветления свекловичный сок в виде сиропа поступает в вакуумный аппарат, где проходит кристаллизацию, и идёт в утфель, эта смесь которая состоит из 50% сахарных кристаллов и 50% сахарного раствора.
Кристаллический сахар потом сушат, и получают сахарный песок, а раствор же вновь варят в вакуумных аппаратах, чтобы выделить оставшийся сахар. После этого получается жёлтый сахар, который вновь проходит переработку и меласса.
За всем этим процессом и оборудованием контролировала смена Владимира Михайловича Смолькина. В их кабинете перед каждым стоит компьютер, по которому виден весь технологический процесс.
В их кабинете перед каждым стоит компьютер, по которому виден весь технологический процесс.
Готовый сахар поступает в упаковочный цех, а затем на склад готовой продукции.
60 лет — на благо республики и её жителей
В этом году завод отмечает 60-летний юбилей. Он пущен в эксплуатацию в 1961 году, в это же время осенью. Тогда за сутки выпускали 1,5 тысяч тонн сахара, сейчас эти показатели увеличились до 7,5 тысяч тонн. Производительность выросла в 5 раз. Это достигнуто в результате обновления производства. Сегодня в ООО «Ромодновосахар» трудятся около 700 человек. Каждый из них делает большое дело не только для своей республики, но и для всей страны.
http://agro.e-mordovia.ru/
Технология производства сахарной свеклы: что изменилось?
Улучшения в производстве сахарной свеклы включают улучшенную генетику, более точное применение удобрений, сокращение обработки почвы и борьбу с болезнями.
Имея возможность работать в Мичиганском государственном университете в сфере сельскохозяйственного производства более 34 лет, было интересно наблюдать за происходящими технологическими изменениями. За последние 15 лет в производстве сахарной свеклы произошли очень существенные изменения. Новые технологические достижения добавили дополнительные улучшения в эффективности, урожайности и качестве.Производители сахара в Мичигане быстро внедрили новые технологии. В результате за последние семь лет промышленность Мичигана произвела более 18 процентов сахара, а за последние 15 лет урожайность увеличивалась в среднем на 0,5 тонны с акра в год.
Произведены улучшения семян сахарной свеклы , которые включают генетику, обработку семян, обработку и затравку. Безусловно, самым запоминающимся изменением стало введение семян Round-Up Ready, которые произвели революцию в борьбе с сорняками сахарной свеклы.Генетические улучшения урожайности, содержания сахара и устойчивости к болезням и вредителям, включая устойчивость к нематодам, также были значительными.
Способность семеноводческих компаний идентифицировать и обрабатывать высококачественные семена также улучшилась. Все семена теперь гранулированы и загрунтованы, что обеспечивает лучшее расстояние между семенами, быстрое появление всходов и более стабильные насаждения. Было введено несколько новых обработок семян, которые включают системные инсектициды и фунгициды, которые обеспечивают лучший контроль над болезнями рассады.
Удобрение Приложения были доработаны, что позволяет производителям вносить удобрения и известь с переменной нормой.В настоящее время доступны многие продукты азотных удобрений, которые предлагают медленное высвобождение или другие средства для предотвращения потерь. Была показана важность внесения удобрений, и большинство производителей используют стартовое удобрение два на два. Несколько лет назад в основном использовались сухие гранулированные удобрения, а сейчас стали преобладать жидкие.
Методы обработки почвы и севообороты также сильно изменились со временем..files/image024.gif) Осенняя вспашка отвала была обычной практикой, и сахарную свеклу высаживали после культур с низким содержанием остатков, таких как сухие бобы.Это было заменено некоторой формой вспашки или рыхления осенним долотом, когда примерно половина урожая следует за кукурузой.
Осенняя вспашка отвала была обычной практикой, и сахарную свеклу высаживали после культур с низким содержанием остатков, таких как сухие бобы.Это было заменено некоторой формой вспашки или рыхления осенним долотом, когда примерно половина урожая следует за кукурузой.
Усовершенствования посевного оборудования позволили нам успешно сажать сахарную свеклу в стебли кукурузы с большим количеством пожнивных остатков или в покровные культуры. Это не только хорошая практика сохранения почвы, но и значительно снижает вероятность выброса сахарной свеклы. Посев несвежих семян, сокращение вторичной обработки почвы и отказ от культивации стали теперь обычной практикой.
Борьба с болезнями и вредителями сахарной свеклы также резко изменилась.Теперь производители могут использовать компьютерное моделирование BEETcast для более точного опрыскивания для борьбы с пятнистостью листьев Cercospora. Теперь с Rhizoctonia можно бороться с помощью подбора сортов и внесения Quadris в борозды или листвы.
Свекольные цистообразующие нематоды раньше боролись только с помощью длительных ротаций. Сегодня сорта, устойчивые к нематодам, наряду с посадкой масличной редьки в качестве ловушки вернули рентабельность бедным продуктивным полям.
RTK, смартфоны и лесозаготовительное оборудование — все это относительно недавние технологические изменения.Технология RTK позволила производителям точно определять урожайность полей, вносить удобрения, а также обеспечивать работу оборудования с помощью автоматического управления. Интернет и его доступность для смартфонов и планшетов сделали информацию доступной у вас под рукой в любое время дня и ночи. Эта технология произвела революцию в сельском хозяйстве и будет продолжаться в обозримом будущем.
Значительно улучшено все оборудование для уборки сахарной свеклы за счет уменьшения тары и повышения эффективности сбора урожая.Самоходные свеклоуборочные комбайны в настоящее время стали обычным явлением, и сахарную свеклу обычно помещают в бурты в конце поля.
Ожидается, что в будущем технологические усовершенствования в производстве и оборудовании будут происходить устойчивыми темпами. Часто эти улучшения могут принести пользу производству сахарной свеклы лишь небольшими порциями. Однако реализация ряда небольших изменений в сочетании приведет к значительным результатам, которые повысят производительность, рентабельность и эффективность.
Была ли эта статья полезной для вас?
Расскажите, почему
Представлять на рассмотрениеПроизводство сахара – обзор
6.3 НАУЧНО-ИССЛЕДОВАТЕЛЬСКАЯ И ТЕХНОЛОГИЧЕСКАЯ РАЗРАБОТКА В СЕЛЬСКОМ ПРОИЗВОДСТВЕ САХАРНОГО ТРОСТА
Сахарный тростник выращивают в Бразилии в основном для производства сахара и этанола. Для удовлетворения производственных потребностей тростник выращивают с использованием так называемой системы ratoon, при которой первый укос делается через 18 месяцев после посадки, а затем ежегодные укосы в течение 4 или 5 лет с уменьшением урожайности. Теплый бразильский климат с дождливым летом и ясным небом зимой помогает тростнику формировать прочную волокнистую структуру на этапе роста и фиксировать сахар зимой.
Теплый бразильский климат с дождливым летом и ясным небом зимой помогает тростнику формировать прочную волокнистую структуру на этапе роста и фиксировать сахар зимой.
Развитию посевов сахарного тростника в Бразилии способствовали постоянные исследования по выведению новых сортов, в частности, проводимые Агрономическим институтом Кампинаса (IAC), Кооперативом производителей сахара и этанола штата Сан-Паулу (Copersucar) и Национальный план по сахарному тростнику (Planalsucar), подразделение Института сахара и алкоголя (IAA).Однако за последние 30 лет весь сельскохозяйственный исследовательский аппарат Сан-Паулу, включая исследовательские станции, подвергался постоянному демонтажу, что замедлило повышение производительности производства сахарного тростника в то время, когда цены на нефть падали. Это сделало этанол менее конкурентоспособным. Тем временем федеральное правительство проводило политику устранения государственного вмешательства в сахарный бизнес, ликвидировав, среди прочих организаций, IAA и Planalsucar.
Некоторые из ранее исследовательских обязанностей были взяты на себя частными организациями.Copersucar, например, сместила акцент своей программы на разработку новых сортов главным образом через CTC, технологический центр в Пирасикабе, Сан-Паулу. Еще одна постоянная задача в исследованиях связана с продлением сезона измельчения за счет развития раннего созревания и повышения урожайности за счет борьбы с вредителями. Программа селекции сахарного тростника также включает современные методы молекулярной биологии. Генетическая трансформация сортов сахарного тростника была осуществлена в лабораториях CTC, и в настоящее время различные трансгенные сорта сахарного тростника проходят полевые испытания.
Еще одно важное технологическое изменение, наблюдаемое в выращивании сахарного тростника за последние 40 лет, связано с внедрением техники для подготовки и сохранения почвы, особенно в южных штатах Бразилии. В последние два десятилетия наблюдается некоторая интенсификация использования машин на различных работах, от обработки почвы до уборки урожая и особенно при погрузке камыша. Менее значительные успехи были подтверждены в посадке тростника. Весь тростник по-прежнему сажают вручную, хотя недавно компания Copersucar разработала машину для посадки тростника и предоставила DMB лицензию.Sermag и Brastoft также предлагают на рынке посадочные машины.
Менее значительные успехи были подтверждены в посадке тростника. Весь тростник по-прежнему сажают вручную, хотя недавно компания Copersucar разработала машину для посадки тростника и предоставила DMB лицензию.Sermag и Brastoft также предлагают на рынке посадочные машины.
Несмотря на прогресс, достигнутый в производстве тростника, сбор урожая остается наименее развитой операцией. Тростниковые поля теперь систематически сжигают, чтобы можно было собирать урожай вручную. Однако это быстро меняется. Давление окружающей среды, соблюдение законодательства и снижение затрат подталкивают к механическому сбору несгоревшего тростника (Furlani et al., 1996). Кроме того, возможность получения дохода от отходов тростника, вероятно, станет стимулом в этом направлении, как только производители начнут оценивать другие варианты получения дохода в своей производственной цепочке.
Развитие сбора урожая тростника и его потенциал для снижения затрат на сельское хозяйство будут обсуждаться далее в этой главе. Другие возможности снижения затрат связаны с оптимизацией управления сельским хозяйством, включая внедрение методов исследования операций и точного земледелия, что позволит более рационально использовать ресурсы и повысить урожайность. Например, концепция производственной среды сочетает карты почв с климатическими данными и данными о сортах и приносит прибыль около 40 долл. США/га согласно испытаниям, проведенным на некоторых сахарных заводах.
Другие возможности снижения затрат связаны с оптимизацией управления сельским хозяйством, включая внедрение методов исследования операций и точного земледелия, что позволит более рационально использовать ресурсы и повысить урожайность. Например, концепция производственной среды сочетает карты почв с климатическими данными и данными о сортах и приносит прибыль около 40 долл. США/га согласно испытаниям, проведенным на некоторых сахарных заводах.
Информационное программное обеспечение обеспечивает основу для значительных улучшений в сельском хозяйстве. Программное обеспечение на основе ГИС (географическая информационная система) со встроенной электроникой вместе с данными о продуктивности может указать влияние переменных, связанных с продуктивностью, таких как плодородие почвы, вредители, болезни, насекомые, сорняки, уплотнение почвы и методы сбора урожая. Это может служить основой для более эффективного управления посевами и повышения производительности. Также разрабатывается программное обеспечение для улучшения логистики, помогающее улучшить распределение погрузчиков, комбайнов или грузовиков для оптимизации потоков сырья на завод. Оптимизация логистики при обрезке и транспортировке тростника уже привела к сокращению сельскохозяйственных затрат на тростник на 5-13 процентов (данные основаны на заводах Copersucar в Сан-Паулу, около 25 процентов производства Бразилии в 1999 году).
Оптимизация логистики при обрезке и транспортировке тростника уже привела к сокращению сельскохозяйственных затрат на тростник на 5-13 процентов (данные основаны на заводах Copersucar в Сан-Паулу, около 25 процентов производства Бразилии в 1999 году).
Влияние даты уборки сахарной свеклы на ее технологические показатели качества по данным разведочного анализа
Выбор даты уборки является одной из основ марочного качества и хорошего выхода сахара из сахарной свеклы. Однако трудно точно и абсолютно определить дату сбора урожая, а точнее дату созревания корнеплодов свеклы.Действительно, зрелость делится на несколько стадий и степеней в зависимости от экологических и климатических условий, таких как температура, осадки, географический район и другие. В настоящем исследовании с использованием наиболее популярного хемометрического метода было выявлено влияние трех сроков сбора урожая (при оцененной зрелости, через 7 дней после созревания и через 15 дней после созревания) на технологические параметры качества, а именно: сахарозу, азот, калий и натрий. анализ главных компонентов (PCA). Для этого использовались образцы из орошаемого периметра Тадла.Результаты поискового анализа с применением АКП ясно показали важное влияние даты сбора урожая на три параметра качества: состав сахарозы, калия и натрия. Но что касается состава азота, между образцами были незначительные различия.
анализ главных компонентов (PCA). Для этого использовались образцы из орошаемого периметра Тадла.Результаты поискового анализа с применением АКП ясно показали важное влияние даты сбора урожая на три параметра качества: состав сахарозы, калия и натрия. Но что касается состава азота, между образцами были незначительные различия.
1. Введение
Сахарная свекла ( Beta vulgaris L.) – двудольное растение, принадлежащее к семейству Chenopodiaceae, известному сегодня как Amaranthaceae, и отряду Caryophylla, имеющему фотосинтетическую систему C3 [1].Это конический корень, пересеченный двумя сахароносными бороздами и имеющий плоскую шейку; он почти полностью зарыт в землю. Корень сахарной свеклы характеризуется очень высоким содержанием растворимых сахаров, которое зависит от нескольких переменных, а именно от региона, времени года, биологии растений, агрономии, времени сбора урожая и послеуборочной практики [1]. Свекла — двулетнее растение, вегетационный цикл которого длится два года. Процесс производства сахара происходит в течение первого года; в этот период запасы сахара максимальны и сахар, образующийся в листьях в результате фотосинтеза, запасается в виде сахарозы в корне.Это постоянная транслокация сахарозы из листьев, которая запасается в концентрических кольцах сосудистой ткани, происходящей из вторичного камбия, возникающего при раннем развитии корней, и в клетках паренхимы, число которых увеличивается во время роста [2]. Таким образом, в это время проводится сбор урожая. В течение второго года растение размножается; затем он использует свои запасы сахара для производства цветочного стебля, который превращается в плоды и семена, чтобы продолжить свой вегетативный цикл [3].Сахарная свекла — это культура, которую можно сажать как одну из многих севооборотных культур с целью полного использования земли и ресурсов в сельскохозяйственной системе [4]. Кроме того, это очень стабильное растение может быть устойчивым к водному стрессу [5], морозоустойчивым [6] или даже устойчивым к солевому стрессу [7, 8] и может иметь более высокую эффективность использования воды и азота, чем другие технические культуры, такие как кукуруза.
Процесс производства сахара происходит в течение первого года; в этот период запасы сахара максимальны и сахар, образующийся в листьях в результате фотосинтеза, запасается в виде сахарозы в корне.Это постоянная транслокация сахарозы из листьев, которая запасается в концентрических кольцах сосудистой ткани, происходящей из вторичного камбия, возникающего при раннем развитии корней, и в клетках паренхимы, число которых увеличивается во время роста [2]. Таким образом, в это время проводится сбор урожая. В течение второго года растение размножается; затем он использует свои запасы сахара для производства цветочного стебля, который превращается в плоды и семена, чтобы продолжить свой вегетативный цикл [3].Сахарная свекла — это культура, которую можно сажать как одну из многих севооборотных культур с целью полного использования земли и ресурсов в сельскохозяйственной системе [4]. Кроме того, это очень стабильное растение может быть устойчивым к водному стрессу [5], морозоустойчивым [6] или даже устойчивым к солевому стрессу [7, 8] и может иметь более высокую эффективность использования воды и азота, чем другие технические культуры, такие как кукуруза. , пшеница и люцерна [9]. Сахарная свекла выращивается в 41 стране мира на площади 8,1 млн га [8, 10] и уже более века выращивается для повышения выхода и чистоты сахарозы и в настоящее время составляет почти 30% мирового производства сахара [11]. ]; этот вклад в мировое производство также был обнаружен Salim et al.[12]. По данным FAO 2019 [13], классификация первой десятки производителей сахарной свеклы выглядит следующим образом: Россия, Франция, Германия, Соединенные Штаты Америки, Турция, Польша, Китай, Египет, Украина и Великобритания.
, пшеница и люцерна [9]. Сахарная свекла выращивается в 41 стране мира на площади 8,1 млн га [8, 10] и уже более века выращивается для повышения выхода и чистоты сахарозы и в настоящее время составляет почти 30% мирового производства сахара [11]. ]; этот вклад в мировое производство также был обнаружен Salim et al.[12]. По данным FAO 2019 [13], классификация первой десятки производителей сахарной свеклы выглядит следующим образом: Россия, Франция, Германия, Соединенные Штаты Америки, Турция, Польша, Китай, Египет, Украина и Великобритания.
На национальном уровне (Марокко) площадь посевов сахарной свеклы увеличилась примерно на 4% и достигла примерно 61 000 га во время кампании 2015/2016 гг. по сравнению с 58 000 га в предыдущей кампании, а общий объем производства увеличился примерно до 61 000 га в течение 2015/2016 гг. кампании и достигла 4,2 млн т, увеличившись на 17 % по сравнению с производством прошлого года [14], а средняя урожайность сахарной свеклы выросла с 61 до 71 т/га, увеличившись на 16 % по сравнению с предыдущей кампанией. [14].На региональном уровне средний объем производства, полученный в периметре Тадла в рамках кампании 2015/2016 гг., составляет около 813 000 т или 26% от достижения. Посевная площадь составляет около 13 040 га в 2015/2016 гг. против 11 200 га в 2009/2010 гг. [15].
[14].На региональном уровне средний объем производства, полученный в периметре Тадла в рамках кампании 2015/2016 гг., составляет около 813 000 т или 26% от достижения. Посевная площадь составляет около 13 040 га в 2015/2016 гг. против 11 200 га в 2009/2010 гг. [15].
В Марокко сахарная свекла – это осенняя культура, которую сеют в октябре и ноябре и собирают в июне, июле и августе. Срок уборки свеклы не определяется стадией физиологической зрелости, а убирают эту культуру, когда ее сахарообразование является оптимальным.Зрелость свеклы, приводящая к пожелтению листьев, трудно точно оценить. Исследования, проведенные в различных марокканских свекловодческих районах, показывают, что фаза созревания свеклы должна быть максимально солнечной и достаточно продолжительной, однако без преувеличения [16]. В целом содержание сахара в корнеплоде имеет колоколообразную кривую: в апреле-начале мая оно слишком низкое, в конце мая приемлемое, в июне хорошее, в июле высокое, в августе снижается [16, 17].
Сразу после уборки свекла проходит различные этапы перед отправкой на сахарный завод (очистка листьев, переворачивание и погрузка в грузовики), поэтому время между уборкой урожая и переработкой на заводе должно быть коротким. чтобы сохранить их высокое содержание сахара, потому что после извлечения из земли содержание сахара в них быстро снижается.Влияние изучения сроков сбора урожая на сельскохозяйственные культуры является бичом, касающимся не только сахарной свеклы, но и других фруктов и овощей, и этим вопросом занимались многие исследователи. Надори и др. [18] изучали влияние подвоя и даты сбора урожая на качество плодов мандарина при хранении в холодильнике; и Shid [20], со своей стороны, изучали влияние сезона сбора урожая на летучие компоненты аромата двух сортов бананов, Feyem et al.[21] изучали влияние даты сбора урожая на прорастание семян некоторых сортов богарного риса, и, наконец, Dibi et al. [22] определили период сбора урожая сортов сладкого картофеля.
Именно в связи с этим была проведена данная работа с целью изучения влияния сроков уборки сахарной свеклы, а именно с опозданием на 5, 10 и 15 дней, на ее технологическое качество, сахарозу и мелассогенность. элементы. Кроме того, исследование аналитических результатов проб было изучено с применением анализа главных компонентов (АГК) с целью визуализации числовых данных в виде пространственного представления для группировки проб с высоким или низким содержанием в зависимости от урожая. дату и изучаемые параметры.
элементы. Кроме того, исследование аналитических результатов проб было изучено с применением анализа главных компонентов (АГК) с целью визуализации числовых данных в виде пространственного представления для группировки проб с высоким или низким содержанием в зависимости от урожая. дату и изучаемые параметры.
2. Материалы и методы
Настоящее исследование проведено на орошаемом периметре Тадла в сельскохозяйственные сезоны 2011-2012 гг.
2.1. Растительное сырье
Всего было использовано 70 корнеплодов сахарной свеклы (каждые 5 корнеплодов представляют собой исследуемый образец), все с одного участка. Они были собраны в контролируемых условиях. Все образцы принадлежали к одному и тому же моногермичному сорту ARDAN-N со средним содержанием сахарина, средней урожайностью и средним циклом, и все они выращивались в одинаковых условиях; дата посева 3 октября 2011 г., почва суглинистая с рН, близким к нейтральному, способ посева механический, внесение удобрений было разделено на два вида внесения, первое – основное удобрение, которым был Phosphate Diammoniaque 18. .46 или DAP, а вторым входом было покровное удобрение Аммонитрат 33,5%, которое применялось дважды, предшествующей культурой были зерновые, и, наконец, использовался тот же режим орошения, что и самотеком с расходом 20 л/с. Однако никаких фитосанитарных обработок не проводилось.
.46 или DAP, а вторым входом было покровное удобрение Аммонитрат 33,5%, которое применялось дважды, предшествующей культурой были зерновые, и, наконец, использовался тот же режим орошения, что и самотеком с расходом 20 л/с. Однако никаких фитосанитарных обработок не проводилось.
Для изучения влияния даты сбора урожая на качество сахарной свеклы использовали следующий процесс: (i) 25 корнеплодов сахарной свеклы собирают в расчетный день созревания: 30 мая 2012 г. (ii) 20 корнеплодов сахарной свеклы собирают через 7 дней: 06 июня 2012 г. (iii) 25 корнеплодов сахарной свеклы убрано через 15 дней: 14 июня 2012 г.
Анализы проводились в день сбора урожая после подготовки репрезентативных проб.Каждая изучаемая репрезентативная проба представляла собой смесь из 5 корнеплодов свеклы таким образом, чтобы иметь 5 репрезентативных проб (А1, А2, А3, А4 и А5) для даты первого урожая, 4 репрезентативных пробы (Е1, Е2, Е3 и Е4) для первой даты сбора урожая. 2-я дата сбора урожая и, наконец, 5 репрезентативных образцов (J1, J2, J3, J4 и J5) для 3-й даты сбора урожая.
Максимальное время между сбором и анализом проб не более 3 часов.
2.2. Приготовление проб
Свекла выкорчевывается вручную, доброкачественная, неразломанная и очищается вручную в поле после выкорчевывания.Все корнеплоды свеклы промывали, следя за тем, чтобы в осахаривающей борозде не осталось земли, затем срезали корневую шейку. Затем отбирали пробу (образец в виде решетки, вес которой составлял 10% от исходного веса, чтобы можно было легко извлечь растворимые вещества) и, наконец, проводили гомогенизацию.
2.3. Приготовление прозрачного раствора
Образец 40 г гомогенизированного рашпиля помещали на глянцевую бумагу; затем рашпиль переносили в химический стакан цепи автоматического разложения и добавляли 165 мл разбавленного раствора ацетата свинца Pb (CH 3 COO) 2 (3H 2 O).Обычно общий объем составлял 200 мл, и крышку помещали на химический стакан при тщательном встряхивании. Через 15 мин пробы фильтровали.
2.4. Химические анализы
Все анализы проводились в соответствии с процедурами, описанными техническим комитетом по стандартизации сахаров (СНИМА, 2005 г.) [23].
2.4.1. Определение сахарозы
Его проводили с использованием сахариметра, который представляет собой сахариметр-поляриметр (марка Schmidt + Haensch, тип SACCHAROMAT) с делениями, определяющими процентное содержание (%) сахара, содержащегося в решетке, и который был специально разработан для сахарной промышленности.
2.4.2. Определение мелассогенных элементов
Это были элементы, снижающие извлечение сахара, а именно Na, K и N α -амино. Концентрации этих элементов в сахарном соке дают представление о мелассном сахаре; то есть, сколько сахара не может быть извлечено классическим процессом сахарной промышленности. Концентрации выражены в ммоль на 100 г сахарного сока . Определение натрия и калия проводили пламенной фотометрией (тип PHF 104), которая заключается в испарении анализируемого образца в пламени, вызывающем возбуждение атомов. При переходе электронов на более стабильный энергетический уровень испускаются фотоны [24]. Фотометрия заключается в установлении связи между интенсивностью излучения и концентрацией анализируемого элемента. Эмиссии натрия и калия были измерены при 589,6 нм и 766,5 нм соответственно.
При переходе электронов на более стабильный энергетический уровень испускаются фотоны [24]. Фотометрия заключается в установлении связи между интенсивностью излучения и концентрацией анализируемого элемента. Эмиссии натрия и калия были измерены при 589,6 нм и 766,5 нм соответственно.
Для определения азота использовался метод Кррутерса и Олдфилда [25], основанный на колориметрическом методе Мура и Штейна [26]. Он позволяет измерять альфа-аминоазот в свекольном соке после дефекации свинца.Этот метод был официально принят в качестве эталонного метода ICUMSA (Международная комиссия по унифицированным методам анализа сахара).
Образец подвергали действию гидразина (N 2 H 4 ) для денатурации белков. Свободные аминокислоты реагируют с нингидрином с образованием комплекса синего цвета в присутствии пропанола. Затем измеряли оптическую плотность этого раствора при 570 нм.
2.5. Анализ основных компонентов
PCA считается основным методом анализа многомерных данных, когда все наблюдаемые переменные имеют числовой тип, и выявления каких-либо связей между этими переменными и между выборками. Его целью является описание данных, содержащихся в таблице с n строк (индивидов/выборок) и столбцов (переменных/параметров) [27].
Его целью является описание данных, содержащихся в таблице с n строк (индивидов/выборок) и столбцов (переменных/параметров) [27].
Главные компоненты получены путем диагонализации матрицы двумерных корреляций. Эта диагонализация определяет набор собственных значений, наблюдение которых для каждой компоненты позволяет определить количество исследуемых графов [28]. Заключительный этап PCA состоит из графического представления, которое обеспечивает обзор результатов, которые не дают численные выражения [29].
Статистический анализ различных аналитических данных был выполнен с использованием программного обеспечения Unscrambler версии 10.2.
3. Результаты и обсуждение
3.1. Однофакторное описательное исследование показателей качества сахарной свеклы в три срока уборки
Результаты различных аналитических и статистических данных, полученных в течение трех сроков уборки, обобщены в таблице 1 и представлены в виде графика на рисунке 1.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

 09
09 65
65  65
65
по сравнению с таблицей 1, сравнение среднего и медиана каждой переменной (изученный параметр) показывает, что распределение каждой переменной было асимметричным, что предполагает наличие выбросов по отношению к каждой из переменных.Но с многомерной точки зрения (при одновременном рассмотрении всех параметров) эти значения не могут быть выбросами. Таким образом, диапазон (максимум-минимум) и стандартное отклонение каждой переменной в три даты сбора урожая были относительно высокими. Позже это может объяснить большой разброс выборок по каждой переменной (см. рис. 1).
Позже это может объяснить большой разброс выборок по каждой переменной (см. рис. 1).
Это одномерное описательное исследование переменных в необработанном состоянии позволило заподозрить наличие выбросов. Кроме того, это позволило обосновать применение PCA на центрально-редуцированных данных.То есть проводится многофакторный описательный анализ, не зависящий от шкал измерения изучаемых параметров.
Это одномерное описательное исследование переменных в необработанном состоянии позволило заподозрить наличие выбросов. Кроме того, это позволило обосновать применение PCA на центрально-редуцированных данных. То есть проводится многофакторный описательный анализ, не зависящий от шкал измерения изучаемых параметров.
3.2.Описательный анализ параметров качества сахарной свеклы в трех датах сбора по ACP
Цель анализа основных компонентов (или PCA) является чисто описательной: это вопрос «исследования» набора наблюдений, собранных в виде таблицы данных. с указанием для каждой статистической единицы (здесь — выборки свеклы) наблюдаемых значений определенного количества количественных переменных (здесь — изучаемых показателей качества).
Анализ главных компонентов является одним из наиболее важных методов многомерного анализа.Это позволяет хорошо визуализировать структуры, присутствующие в данных. На самом деле, интерпретация результатов PCA обычно обеспечивается визуализацией оценок графиков и графиков нагрузок [30]. Балл выборки — это значение на оси главных компонентов, куда оно проецируется [31]. Нагрузки исходных переменных можно интерпретировать как корреляции между этими переменными и главными компонентами. Чем больше вклад (нагрузка) переменной на главный компонент, тем больше переменная связана с этим компонентом.Вкратце, график нагрузок позволяет определить и определить наиболее важные переменные в соответствии с их распределением по главным компонентам.
При использовании процедуры PCA количество исходных переменных сокращается до нескольких главных компонентов (PC), которые по-прежнему представляют основную информацию исходного набора данных. ПК выбираются таким образом, что первый ПК несет большую часть информации, за ним следует второй ПК, несущий меньше информации, и так далее в порядке убывания. После определенного количества ПК изменение, смоделированное новым ПК, может быть связано в основном с шумом.
После определенного количества ПК изменение, смоделированное новым ПК, может быть связано в основном с шумом.
После импорта данных в программу Unscrambler сначала к необработанным аналитическим данным был применен анализ основных компонентов (PCA). Затем для работы с корреляционной матрицей была рассчитана редуцированная по центру матрица данных, чтобы описательный анализ не зависел от единиц измерения.
Таким образом, анализ основных компонентов (PCA) был выполнен на основе аналитических данных (необработанных и центрально-обведенных) из 14 образцов свеклы, где интерпретация результатов основана на визуализации двойного графика (рис. 2) и корреляционных нагрузках (рис. 3). ).
Рисунок 2 (представление выборок/переменных) представляет собой графическое представление, в котором проекция переменных и выборок совмещена на оси ПК. Это представление позволяет визуализировать выборки, имеющие высокие (или низкие) значения данной переменной. Как правило, существует группировка образцов по дате сбора урожая. Согласно рисунку 2(а), эта группировка тесно связана с составом сахарозы (богатством). Хотя это, соответственно, связано с составом калия, сахарозы и натрия (см. рис. 2(b)), эти результаты были подтверждены графиком «корреляционных нагрузок» (рис. 3).
Согласно рисунку 2(а), эта группировка тесно связана с составом сахарозы (богатством). Хотя это, соответственно, связано с составом калия, сахарозы и натрия (см. рис. 2(b)), эти результаты были подтверждены графиком «корреляционных нагрузок» (рис. 3).
На рис. 3(а) представлены одномерные корреляционные нагрузки. Красные пунктирные линии обозначают границы 50- и 100-процентной дисперсии для данной главной компоненты. Переменные, лежащие в верхней и нижней границах графика, моделируются этим основным компонентом. Между двумя внутренними клеммами нет. При интерпретации этой цифры состав сахарозы (богатство) был наиболее важной переменной. Таким образом, он содержал очень большую вариацию; кроме того, он отвечал за группировку образцов по дате сбора урожая.
Чтобы иметь независимую интерпретацию эффекта масштаба между переменными, аналитические данные были сведены к центру. Рисунок 3(b) представляет собой круг корреляции, определяемый двумя основными компонентами. Это графическое представление переменных в соответствии с их коэффициентами корреляции с главными компонентами. Все параметры были расположены в радиусе между двумя эллипсами, что объясняет более 50% и менее 100% дисперсии данных. Итак, четыре изучаемых параметра содержат достаточно структурированную вариацию, позволяющую различать образцы свеклы по дате уборки.
Все параметры были расположены в радиусе между двумя эллипсами, что объясняет более 50% и менее 100% дисперсии данных. Итак, четыре изучаемых параметра содержат достаточно структурированную вариацию, позволяющую различать образцы свеклы по дате уборки.
В данном исследовании срок уборки доказал свое влияние на различные качественные показатели сахарной свеклы, что также подтверждается библиографией. Более того, есть исследователи, изучавшие влияние различных сроков уборки на урожайность и технологичность сахарной свеклы в зависимости от засоренности и подмерзания корнеплодов [32] и обнаружившие зависимость от урожайности и качества урожая. корнеплодов свеклы (калий, натрий, альфа-аминоазот, сахар) и срок сбора урожая, а также оценивали степень зависимости.Другие исследователи изучали влияние урожая на потери при хранении [33] и показали, что качество урожая имеет решающее значение для дальнейшей оптимизации цепочки создания стоимости. Другие полагались на дроны-коллекторы мультиспектральных изображений, способные определять вегетационный индекс для оценки сроков сбора урожая сахарной свеклы [34]; они обнаружили, что ранние сроки сбора урожая снижают выход извлекаемого сахара по сравнению с поздними сроками сбора урожая. В другом исследовании изучалось взаимодействие между датой посева и генотипами сахарной свеклы для разных сроков сбора урожая [35]; результаты анализа показали, что генотипы, дата посадки и дата сбора урожая значительно повлияли на выход сахара.
В другом исследовании изучалось взаимодействие между датой посева и генотипами сахарной свеклы для разных сроков сбора урожая [35]; результаты анализа показали, что генотипы, дата посадки и дата сбора урожая значительно повлияли на выход сахара.
4. Выводы
Сахарная свекла, выращиваемая из-за ее мясистого корня, который формируется в первый год сельскохозяйственного цикла, в основном используется для производства сахара. В сахарной промышленности общее качество свеклы оценивается путем измерения следующих соединений: сахароза, общий азот, натрий и калий. В настоящем исследовании удалось продемонстрировать влияние даты уборки урожая на показатели качества сахарной свеклы. Продление сроков сбора урожая, семь и пятнадцать дней после предполагаемой зрелости, позволило сказать, что имеется значительное влияние, в частности, на содержание сахарозы и двух мелассогенных элементов калия и натрия, в то время как состав азота показывает незначительные изменения. .
Доступность данных
Используемые данные можно получить у первого автора (lamia. [email protected]) по запросу.
[email protected]) по запросу.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Благодарности
Авторы благодарят SUTA за финансирование проекта.
OMICS Технологии и приложения в сахарной свекле
Front Plant Sci. 2016; 7: 900.
Yongxue Zhang
1 Ключевая лаборатория молекулярной биологии провинции Хэйлунцзян, Колледж наук о жизни Хэйлунцзянского университета, Харбин, Китай
2 Инженерно-исследовательский центр технологий сельскохозяйственной микробиологии, Министерство образования, Хэйлунцзянский университет, Харбин, Китай
Jingdong Nan
1 Ключевая лаборатория молекулярной биологии провинции Хэйлунцзян, Колледж наук о жизни Хэйлунцзянского университета, Харбин, Китай
2 Центр инженерных исследований сельскохозяйственной микробиологии, Министерство Образование, Хэйлунцзянский университет, Харбин, Китай
Бин Юй
1 Ключевая лаборатория молекулярной биологии провинции Хэйлунцзян, Колледж наук о жизни Хэйлунцзянского университета, Харбин, Китай
2 Инженерно-исследовательский центр технологий сельскохозяйственной микробиологии, Министерство образования, Хэйлунцзян У niversity, Харбин, Китай
1 Ключевая лаборатория молекулярной биологии провинции Хэйлунцзян, Колледж наук о жизни, Хэйлунцзянский университет, Харбин, Китай
2 Инженерно-исследовательский центр технологий сельскохозяйственной микробиологии, Министерство образования, Хэйлунцзянский университет, Harbin, China
Под редакцией: Pingfang Yang, Китайская академия наук, China
Рецензирование: Sixue Chen, Университет Флориды, США; Jingkui Tian, Zhejiang University, China
Эта статья была отправлена в Plant Proteomics, раздел журнала Frontiers in Plant Science
Поступила в редакцию 31 марта 2016 г.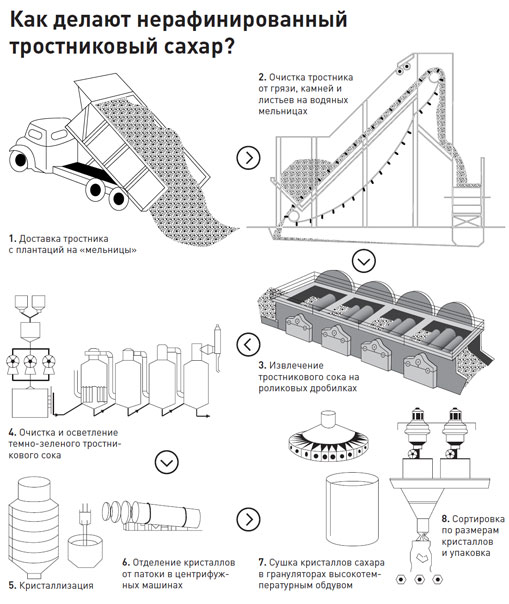 ; Принято 7 июня 2016 г.
; Принято 7 июня 2016 г.
Это статья с открытым доступом, распространяемая на условиях лицензии Creative Commons Attribution License (CC BY). Использование, распространение или воспроизведение на других форумах разрешено при условии указания автора(ов) или лицензиара оригинала и ссылки на оригинальную публикацию в этом журнале в соответствии с принятой академической практикой. Запрещается использование, распространение или воспроизведение без соблюдения этих условий.
Эта статья была процитирована другими статьями в PMC.Abstract
Сахарная свекла относится к семейству Chenopodiaceae . Это важная сахарная культура, которая поставляет примерно 35% сахара в мире. Линия сахарной свеклы М14 представляет собой уникальную зародышевую плазму, содержащую генетические материалы Beta vulgaris L. и Beta corolliflora Zoss. И проявляет толерантность к солевому стрессу. В этом обзоре мы обобщили технологии и приложения OMICS для сахарной свеклы, включая M14, для идентификации новых генов, белков, связанных с биотическими и абиотическими стрессами, апомиксов и метаболитов, связанных с энергией и пищей. Обзор OMICS по открытию новых генов, белков и метаболитов в сахарной свекле помог нам понять сложные механизмы, лежащие в основе многих процессов, таких как апомиксы, устойчивость к биотическим и абиотическим стрессам. Полученные знания ценны для повышения устойчивости сахарной свеклы и других культур к биотическим и абиотическим стрессам, а также для повышения урожайности сахарной свеклы для производства энергии и продуктов питания.
Обзор OMICS по открытию новых генов, белков и метаболитов в сахарной свекле помог нам понять сложные механизмы, лежащие в основе многих процессов, таких как апомиксы, устойчивость к биотическим и абиотическим стрессам. Полученные знания ценны для повышения устойчивости сахарной свеклы и других культур к биотическим и абиотическим стрессам, а также для повышения урожайности сахарной свеклы для производства энергии и продуктов питания.
Ключевые слова: сахарная свекла, геномика, транскриптомика, протеомика, метаболомика
Введение
Сахарная свекла ( Beta vulgaris .L), вид семейства Chenopodiaceae , является важной сахарной культурой, которая поставляет примерно 35% сахара в мире (Liu et al., 2008). В Соединенных Штатах сахарная свекла обеспечивает около 55 процентов всего сахара, производимого внутри страны с середины 1990-х годов (Benoit et al., 2015). Сахарная свекла была завезена в Китай из Аравии около 1500 лет назад и является двудольным растением, имеющим высокую экономическую ценность во многих странах.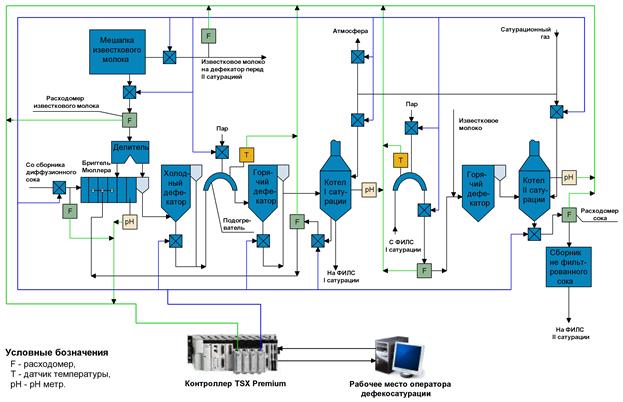 Поэтому вопрос о том, как эффективно выращивать урожай, был приоритетным и широко исследовался (Draycott, 2006).Сахарная свекла — это двулетняя культура, у которой в первый год (стадия вегетации) вырастает стержневой корень, богатый сахаром, а на второй год (стадия репродуктивности) — цветущий семенной стебель; Chen et al., 2016). Виды сахарной свеклы можно различать по различным внутренним и внешним признакам, таким как хозяйственные признаки, разнообразие признаков, хромосомная плоидность. Beta corolliflora Зосс. (2 n = 36) — дикий вид секции свеклы Corollinae, обладающий многими характеристиками, включая устойчивость к засухе, холоду, соли и болезням.Сахарная свекла ( Beta vulgaris ) относится к солеустойчивым культурам (Дунайска-Ордак и др., 2014). Ученые изучали межвидовое скрещивание культурной сахарной свеклы ( Beta vulgaris L. 2 n = 19) и B. corolliflora Zoss. на протяжении десятилетий (Dalke et al., 1971; Filutowicz and Dalke, 1976).
Поэтому вопрос о том, как эффективно выращивать урожай, был приоритетным и широко исследовался (Draycott, 2006).Сахарная свекла — это двулетняя культура, у которой в первый год (стадия вегетации) вырастает стержневой корень, богатый сахаром, а на второй год (стадия репродуктивности) — цветущий семенной стебель; Chen et al., 2016). Виды сахарной свеклы можно различать по различным внутренним и внешним признакам, таким как хозяйственные признаки, разнообразие признаков, хромосомная плоидность. Beta corolliflora Зосс. (2 n = 36) — дикий вид секции свеклы Corollinae, обладающий многими характеристиками, включая устойчивость к засухе, холоду, соли и болезням.Сахарная свекла ( Beta vulgaris ) относится к солеустойчивым культурам (Дунайска-Ордак и др., 2014). Ученые изучали межвидовое скрещивание культурной сахарной свеклы ( Beta vulgaris L. 2 n = 19) и B. corolliflora Zoss. на протяжении десятилетий (Dalke et al., 1971; Filutowicz and Dalke, 1976). В нашей лаборатории Guo et al. получили моносомную аддитивную линию сахарной свеклы М14 (рис. ), которая содержит геном Beta vulgaris L. с добавлением No.9 хромосома B. corolliflora Zoss (Guo et al., 1994). Он имеет несколько интересных характеристик, включая апомиксы и устойчивость к засухе, холоду и солевому стрессу (Guo et al., 1994). Апомиксис — это способ бесполого размножения, характеризующийся образованием клональных семян посредством развития партеногенеза нередуцированного яйца. Апомиктический процесс обходит мейоз и оплодотворение яйцеклетки, производя потомство, которое является точной копией материнского растения (Nogler, 1984; Ozias-Akins, 2006).Таким образом, сахарная свекла М14 может служить уникальной зародышевой плазмой для изучения характеристик апомиксов и устойчивости к абиотическим стрессам. .
В нашей лаборатории Guo et al. получили моносомную аддитивную линию сахарной свеклы М14 (рис. ), которая содержит геном Beta vulgaris L. с добавлением No.9 хромосома B. corolliflora Zoss (Guo et al., 1994). Он имеет несколько интересных характеристик, включая апомиксы и устойчивость к засухе, холоду и солевому стрессу (Guo et al., 1994). Апомиксис — это способ бесполого размножения, характеризующийся образованием клональных семян посредством развития партеногенеза нередуцированного яйца. Апомиктический процесс обходит мейоз и оплодотворение яйцеклетки, производя потомство, которое является точной копией материнского растения (Nogler, 1984; Ozias-Akins, 2006).Таким образом, сахарная свекла М14 может служить уникальной зародышевой плазмой для изучения характеристик апомиксов и устойчивости к абиотическим стрессам. .
В ходе эволюции растения выработали сложные стратегии, которые регулируют биохимическую и физиологическую акклиматизацию, чтобы реагировать на биотический стресс (вирусные, бактериальные, грибковые и оомицетовые инфекции; Baum et al.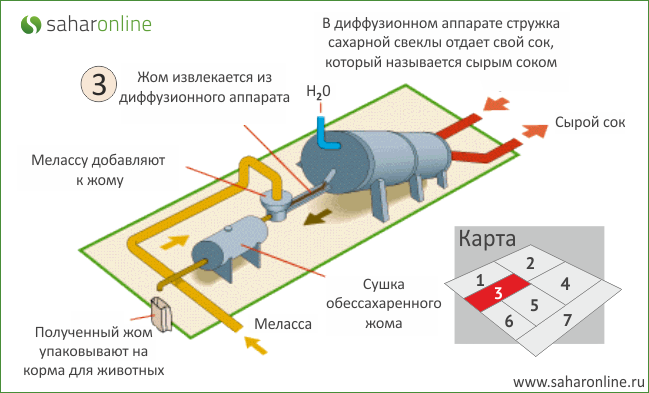 , 2007) и абиотический стресс (соленость, засуха и низкая температура; Barnabas et al., 2008; Yolcu et al., 2016). Биотические и абиотические стрессы серьезно снижают продуктивность сельского хозяйства во всем мире (Munns and Tester, 2008; Pinhero et al., 2011; Mishra et al., 2012). Поэтому понимание того, как растения реагируют и переносят биотические и абиотические стрессы, важно для повышения продуктивности растений (например, сахарной свеклы) в этих сложных условиях. Чтобы свести к минимуму негативное влияние этих стрессов, изучение того, как у сахарной свеклы развились механизмы преодоления стресса, даст новые знания и приведет к новым стратегиям улучшения селекции устойчивой к стрессу сахарной свеклы и других культур.
, 2007) и абиотический стресс (соленость, засуха и низкая температура; Barnabas et al., 2008; Yolcu et al., 2016). Биотические и абиотические стрессы серьезно снижают продуктивность сельского хозяйства во всем мире (Munns and Tester, 2008; Pinhero et al., 2011; Mishra et al., 2012). Поэтому понимание того, как растения реагируют и переносят биотические и абиотические стрессы, важно для повышения продуктивности растений (например, сахарной свеклы) в этих сложных условиях. Чтобы свести к минимуму негативное влияние этих стрессов, изучение того, как у сахарной свеклы развились механизмы преодоления стресса, даст новые знания и приведет к новым стратегиям улучшения селекции устойчивой к стрессу сахарной свеклы и других культур.
В последние годы знания в области геномики, основанные на секвенировании следующего поколения (NGS), системах редактирования генов, подавлении генов и методах сверхэкспрессии, предоставили большой объем генетической информации, помогающей раскрыть механизмы биотических и абиотических стрессовых реакций у растений. (Саад и др., 2013; Шан и др., 2013; Инь и др., 2014; Луан и др., 2015). На уровне транскриптома технологические инновации позволили получить обзор изменений, происходящих на уровне транскриптома при различных стрессовых условиях окружающей среды.Микрочипы и методы секвенирования РНК используются для выяснения дифференциальной экспрессии генов, участвующих в реакциях на биотический и абиотический стресс у различных видов растений (Kreps et al., 2002; Shinozaki and Yamaguchi, 2007; Ergen and Budak, 2009; Mitchell et al. ., 2014; Акпинар и др., 2015; Будак и Акпинар, 2015; Ван и др., 2016). Протеомика и метаболомика — это два новых «омических» метода в постгеномную эпоху (Fernandez-Garcia et al., 2011). Технологии протеомики позволяют одновременно идентифицировать и количественно определять тысячи белков, что является важным инструментом для понимания биологических систем и их регуляции (Silva-Sanchez et al., 2015). Протеомику можно использовать для сравнения протеомов в различных стрессовых условиях (Draycott, 2006; Liu et al.
(Саад и др., 2013; Шан и др., 2013; Инь и др., 2014; Луан и др., 2015). На уровне транскриптома технологические инновации позволили получить обзор изменений, происходящих на уровне транскриптома при различных стрессовых условиях окружающей среды.Микрочипы и методы секвенирования РНК используются для выяснения дифференциальной экспрессии генов, участвующих в реакциях на биотический и абиотический стресс у различных видов растений (Kreps et al., 2002; Shinozaki and Yamaguchi, 2007; Ergen and Budak, 2009; Mitchell et al. ., 2014; Акпинар и др., 2015; Будак и Акпинар, 2015; Ван и др., 2016). Протеомика и метаболомика — это два новых «омических» метода в постгеномную эпоху (Fernandez-Garcia et al., 2011). Технологии протеомики позволяют одновременно идентифицировать и количественно определять тысячи белков, что является важным инструментом для понимания биологических систем и их регуляции (Silva-Sanchez et al., 2015). Протеомику можно использовать для сравнения протеомов в различных стрессовых условиях (Draycott, 2006; Liu et al. , 2008; Benoit et al., 2015). Метаболомика фокусируется на глобальном профиле низкомолекулярных (< 1000 Да) метаболитов, которые являются конечными продуктами метаболизма в биологических жидкостях, тканях и даже во всем организме (Brosché et al., 2005). Метаболомика в последнее время используется во все большем количестве приложений для изучения реакции растительных метаболитов на абиотические стрессы, особенно на засуху, наводнения, засоление и экстремальные температуры (жару и холод; Jorge et al., 2015; Цзя и др., 2016). Очевидно, что комбинация методов OMICS, включая геномику, транскриптомику, протеомику и метаболомику, может служить для проверки и дополнения друг друга, чтобы обеспечить эффективный способ, способный улучшить устойчивость растений к стрессу.
, 2008; Benoit et al., 2015). Метаболомика фокусируется на глобальном профиле низкомолекулярных (< 1000 Да) метаболитов, которые являются конечными продуктами метаболизма в биологических жидкостях, тканях и даже во всем организме (Brosché et al., 2005). Метаболомика в последнее время используется во все большем количестве приложений для изучения реакции растительных метаболитов на абиотические стрессы, особенно на засуху, наводнения, засоление и экстремальные температуры (жару и холод; Jorge et al., 2015; Цзя и др., 2016). Очевидно, что комбинация методов OMICS, включая геномику, транскриптомику, протеомику и метаболомику, может служить для проверки и дополнения друг друга, чтобы обеспечить эффективный способ, способный улучшить устойчивость растений к стрессу.
Сахарная свекла является хорошим растительным ресурсом для изучения и идентификации генов и белков, участвующих в устойчивости к стрессу. Сахарная свекла широко используется в сахарной промышленности (Liu et al., 2008). Это источник чистой энергии с помощью газообразного водорода и биоэтанола (Dhar et al. , 2015). Он содержит большое количество метаболитов бетаина и беталаина. Бетаин используется для повышения стрессоустойчивости растений (Catusse et al., 2008). Беталаины представляют собой природные пигменты, обладающие потенциальной пользой для здоровья (антиканцерогенные и антиоксидантные свойства) и привлекающие как научный, так и экономический интерес (Stintzing and Carle, 2007; Moreno et al., 2008). Богатый и дешевый источник беталаинов в корнеплоде красной свеклы ( Beta vulgaris L.) очень привлекателен для фармацевтической и пищевой промышленности (Wybraniec, 2005; Wybraniec et al., 2011, 2013). В этом обзоре мы обобщили применение OMICS и рассмотрели недавние открытия в исследованиях сахарной свеклы, включая M14 для идентификации новых генов и белков, связанных с биотическими и абиотическими стрессами, апомиксами и метаболитами, связанными с производством энергии и продуктов питания. Полученные знания ценны для повышения устойчивости сахарной свеклы и других культур к биотическим и абиотическим стрессам, а также для повышения урожайности сахарной свеклы для производства энергии и продуктов питания.
, 2015). Он содержит большое количество метаболитов бетаина и беталаина. Бетаин используется для повышения стрессоустойчивости растений (Catusse et al., 2008). Беталаины представляют собой природные пигменты, обладающие потенциальной пользой для здоровья (антиканцерогенные и антиоксидантные свойства) и привлекающие как научный, так и экономический интерес (Stintzing and Carle, 2007; Moreno et al., 2008). Богатый и дешевый источник беталаинов в корнеплоде красной свеклы ( Beta vulgaris L.) очень привлекателен для фармацевтической и пищевой промышленности (Wybraniec, 2005; Wybraniec et al., 2011, 2013). В этом обзоре мы обобщили применение OMICS и рассмотрели недавние открытия в исследованиях сахарной свеклы, включая M14 для идентификации новых генов и белков, связанных с биотическими и абиотическими стрессами, апомиксами и метаболитами, связанными с производством энергии и продуктов питания. Полученные знания ценны для повышения устойчивости сахарной свеклы и других культур к биотическим и абиотическим стрессам, а также для повышения урожайности сахарной свеклы для производства энергии и продуктов питания.
Обзор OMICS для обнаружения новых генов, белков и метаболитов в сахарной свекле
В последние годы значительно возросло использование инструментов OMICS для изучения биотических и абиотических стрессов у растений.Существующие методы включают геномику, транскриптомику, протеомику, метаболомику и ряд других, способных обнаруживать и характеризовать экспрессию генов или белков во время биотических и абиотических стрессов с высокой эффективностью, как показано на рисунке. Эти высокочувствительные инструменты могут анализировать растительные ткани и помогают лучше понять механизмы толерантности, используемые сахарной свеклой.
Схематическая диаграмма, показывающая поток информации для исследований OMICS по сахарной свекле .
Геномные исследования сахарной свеклы
Полногеномная последовательность сахарной свеклы
Полногеномная последовательность сахарной свеклы описана Dohm et al. (2014). В общей сложности 27 421 ген, кодирующий белок, был предсказан на основе данных транскрипции и аннотирован на основе гомологии последовательностей (Dohm et al. , 2014). По сравнению с другими цветковыми растениями с геномной информацией сахарная свекла имеет небольшое количество генов, кодирующих факторы транскрипции. Было высказано предположение, что сахарная свекла может содержать неизвестные гены, связанные с контролем транскрипции, и что сеть генетических взаимодействий сахарной свеклы могла развиваться уникальным образом по сравнению с другими видами.Используя последовательность генома сахарной свеклы и связанные ресурсы, мы ожидали найти молекулярные механизмы, лежащие в основе регуляции генов и взаимодействия генов с окружающей средой. Кроме того, эта информация может помочь в разработке культур с улучшенным производством сахара и натуральных веществ и сыграть важную роль в будущих исследованиях генома растений.
, 2014). По сравнению с другими цветковыми растениями с геномной информацией сахарная свекла имеет небольшое количество генов, кодирующих факторы транскрипции. Было высказано предположение, что сахарная свекла может содержать неизвестные гены, связанные с контролем транскрипции, и что сеть генетических взаимодействий сахарной свеклы могла развиваться уникальным образом по сравнению с другими видами.Используя последовательность генома сахарной свеклы и связанные ресурсы, мы ожидали найти молекулярные механизмы, лежащие в основе регуляции генов и взаимодействия генов с окружающей средой. Кроме того, эта информация может помочь в разработке культур с улучшенным производством сахара и натуральных веществ и сыграть важную роль в будущих исследованиях генома растений.
SMRT генома хлоропластов сахарной свеклы
SMRT (Single Molecule Real-Time) — это метод секвенирования третьего поколения, который предлагает гораздо большую длину считывания по сравнению с методами NGS.Он хорошо подходит для проектов de novo или изменения последовательности. Он содержит не только риды, происходящие из ядерного генома, но также множество ридов из органелл организма-мишени (Sanger et al., 1977; Liu et al., 2012). Стадерманн и др. описал рабочий процесс для сборки de novo генома хлоропластов сахарной свеклы на основе данных, полученных из набора данных секвенирования SMRT, нацеленного на ядерный геном (Stadermann et al., 2015). Они идентифицировали в общей сложности 114 отдельных генов.Из них 79 генов кодируют мРНК (т.е. белки), 7 кодируют рРНК и 28 являются тРНК. Девять генов расположены в областях инвертированных повторов (IR), которые кодируют 5 мРНК, 1 рРНК и 3 тРНК. По сравнению со сборкой Illumina аннотация показала некоторые отличия из-за изменений в базовых последовательностях.
Он содержит не только риды, происходящие из ядерного генома, но также множество ридов из органелл организма-мишени (Sanger et al., 1977; Liu et al., 2012). Стадерманн и др. описал рабочий процесс для сборки de novo генома хлоропластов сахарной свеклы на основе данных, полученных из набора данных секвенирования SMRT, нацеленного на ядерный геном (Stadermann et al., 2015). Они идентифицировали в общей сложности 114 отдельных генов.Из них 79 генов кодируют мРНК (т.е. белки), 7 кодируют рРНК и 28 являются тРНК. Девять генов расположены в областях инвертированных повторов (IR), которые кодируют 5 мРНК, 1 рРНК и 3 тРНК. По сравнению со сборкой Illumina аннотация показала некоторые отличия из-за изменений в базовых последовательностях.
миРНК, участвующие в устойчивости к абиотическим стрессам
миРНК представляют собой короткие некодирующие РНК длиной 19–23 нуклеотида, которые играют регуляторную роль во многих процессах (Budak et al., 2015). miRNAs могут действовать как на транскрипционном, так и на посттранскрипционном уровнях. Опосредованный miRNA механизм молчания генов регулирует экспрессию факторов транскрипции, фитогормонов и др. онтогенетических сигнальных путей (Llave et al., 2002; Dalmay, 2006; Sunkar et al., 2007).
Опосредованный miRNA механизм молчания генов регулирует экспрессию факторов транскрипции, фитогормонов и др. онтогенетических сигнальных путей (Llave et al., 2002; Dalmay, 2006; Sunkar et al., 2007).
Более ранние исследования показали, что миРНК в основном нацелены на факторы транскрипции, но недавние исследования показали, что миРНК также нацелены на другие пути передачи сигналов развития/стресс, которые участвуют в различных физиологических процессах, включая рост и развитие корней, реакцию на стресс, передачу сигнала, морфогенез листа, защита растений и биогенез мкРНК (Curaba et al., 2014). Ли Дж. Л. и соавт. (2015) сообщили о 13 зрелых микроРНК из 12 семейств с использованием подхода in silico , основанного на 29857 тегах экспрессированных последовательностей и 279223 последовательностях обзора генома в B. vulgaris . Целевой сервер psRNA предсказал 25 генов-мишеней для 13 микроРНК. Гены-мишени, показанные в таблице, по-видимому, кодируют факторы транскрипции или участвуют в метаболизме, передаче сигналов, реакции на стресс, росте и развитии.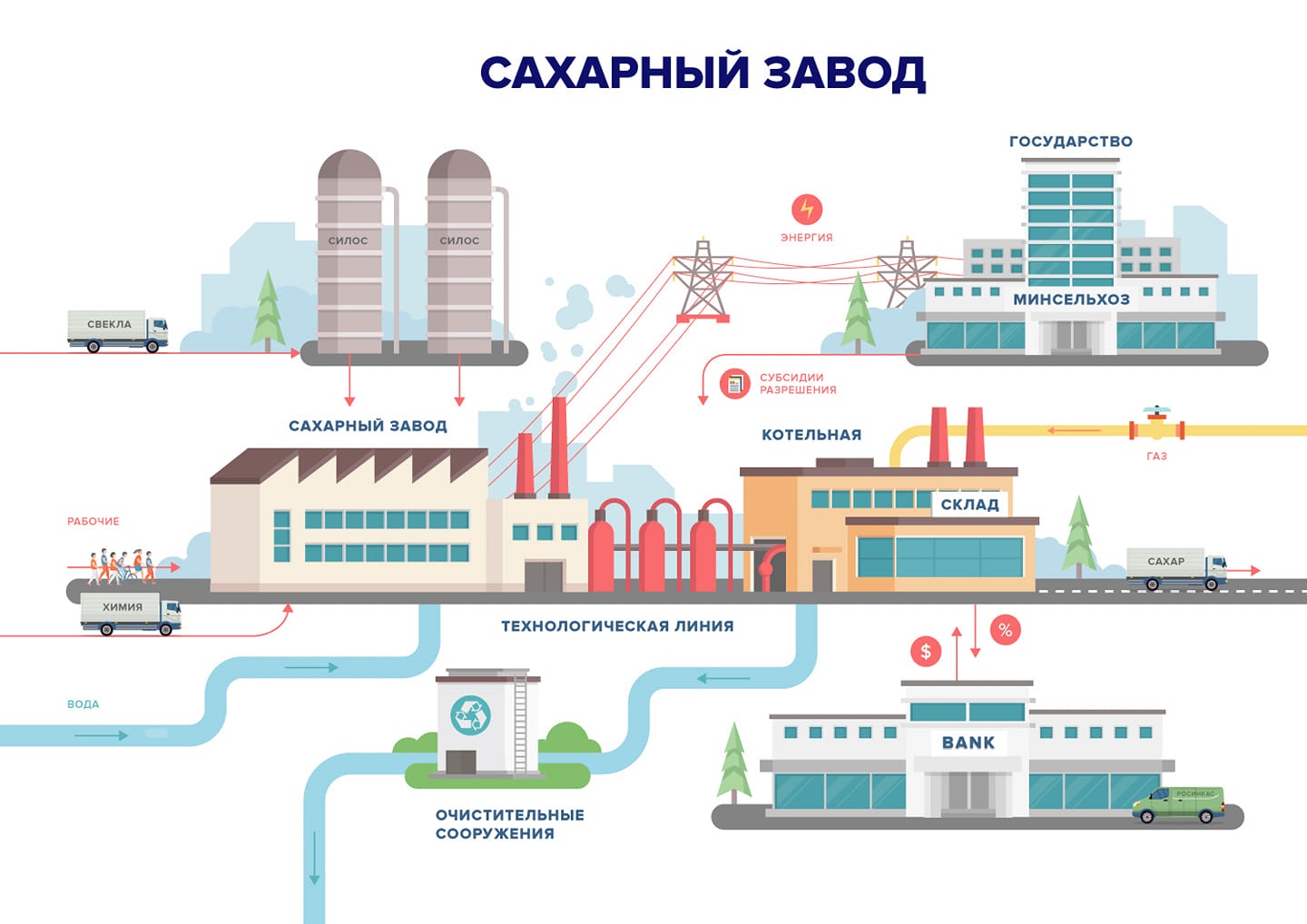 Однако из текущей базы данных сахарной свеклы не было предсказано целей для Bvu-miR4, Bvu-miR9, Bvu-miR10, Bvu-miR11 и Bvu-miR12 .
Однако из текущей базы данных сахарной свеклы не было предсказано целей для Bvu-miR4, Bvu-miR9, Bvu-miR10, Bvu-miR11 и Bvu-miR12 .
Таблица 1
Предсказанные функции микроРНК сахарной свеклы в развитии растений, передаче сигналов и реакциях на стресс .
| Название целевого белка | Go Go (молекулярная функция) | 120991| | | BVU-MIR1 | Белковый белковый белок | Корневая разработка | Moody et al. |  , 2012 , 2012 BVU-MIR2 | ATPASE, V0 Комплекс, C Subunit | Гидролазная активность, действующая на кислотные ангидриды, катализатор трансмембраны перемещения веществ | соль | lehr et al., 1999 | Proton-Transporting ATPASE активность, вращательный ME | — | 9012 — — | — | — — | 9012 | ATPASE, V0 Комплекс, D SUBUNIT | Активность транспортировки ионов водорода | ION относится | озеро et al. | 9012 , 2004 , 2004 RanBPM-Alke | Белкового белка | 9069 | — | — | 9063 9012 | 9063 | Абсцизовая кислота, нечувствительна 5 | Специализированная последовательность ДНК. 2013 | BVU-MIR3 | Nucleoside Phosphatase GDA1 / CD39 | Гидролазная активность | — | 9012 — 9012 — BVU-MIR5 | COPA-POV белок | — | — | — 9012 — | BVU-MIR6 | Diphenol Oxidase | Медь ION ОБЕСПЕЧЕНИЕ | — | — | — — | Гидрохинон: Оксидареудукторедуковаяuctuctaze 20169 | – | – | Фоторецептор PHYA4 | – | – | – | Bvu-miR7 | Оксидазоподобный белок респираторного взрыва | Пероксидазная активность | Биотические стрессы и взаимодействия, абиотические взаимодействия. |  Torres и Dangl, 2005 | 9069 | 9012 | 9063 | 9012 | Активность окиследуют (P)H, кислород в качестве акцептора | Bvu-miR8 | Гистонацетилтрансфераза GCN5 | Связывание с белками | Benhamed. |  1.et9 9091 Транскрипция, передача сигналов 9091, 2008 1.et9 9091 Транскрипция, передача сигналов 9091, 2008 9093 9 | ДНК-транскрипция | ДНК — 9012 —2 — | — | Специфика активность фактора транскрипции | – | – | MYB6 | связывание ДНК | транскрипция, передача сигналов | Chang et al. |  , 2013 , 2013 Chromatin Binding | 9093 | 9012 | NucleoSome Ассамблея белок 1-подобный белок 2 | — | —2 — 9012 — | MCM Белковый белок | ATP Binding | — | — | 9012 9012 | — | Волновой комплекс Scar2 | Actin Bearing | — | 9063 Белок, связывающий элемент, чувствительный к обезвоживанию 1 | ДНК-связывающий | Транскрипция, передача сигналов | Sun et al. |  , 2008 , 20083612 9093 9 | Sequence-специфичные ДНК, привязки транскрипции фактор | BVU-MIR13 | фактор транскрипции WD-Repeat белок 1 | Белковый белок | напряжение | Li et al ., +2014 | тетрапирролов биосинтез | — | — | — | гидроксиметилбилан синтазы | гидроксиметилбилан синтазы | — | — | глицерол-3-фосфат O-ACYLTRANSFERASE | — | — | — | — | Глицерин-3-фосфат O-Acyltransferase Activity | — | — | 9012 | Детализация для передачи, передача ацильных групп | – | – | натрий / сульфатный симпортер | Транспортирован | — | — | Cupredoxin | Медь ионов | — | — | 9012 | 9069 | — | — | — | Удлиненный фактор 1 | 10169 | — | 9069 — | — | 9012 9093 | Reucine, богатый белком, содержащий белок | Белкового белка | Сигнал трансдукции | Гоу и др. |  , 2010 , 2010 Magnesium-Protoporphyrin IX MonomeThyal | — | — | — 9012 9012 | Accidoodooductase Activity | — | — | 9063 | ION ION | — | — | — | 9012 — 9012 — — | |
|---|
Несколько выявленных MIRNAS имеют критические роли в растениях. Например, экспрессия Bvu-miR1 (белок ARABIDILLO) в A. thaliana регулирует развитие многоклеточных корней (Moody et al., 2012). Bvu-miR2 регулирует экспрессию АТФазы во время развития растений и координирует ее индукцию в ответ на сильное засоление (Lehr et al., 1999). Посредством регуляции транскрипции он также влияет на АТФазную активность субъединицы I хелатазы магния в ячмене (Lake et al., 2004) и на нечувствительность к абсцизовой кислоте 5, необходимую для задержки роста проросших проростков в условиях стресса окружающей среды (Liu and Stone, 2013). Bvu-miR7 нацелен на семейство генов респираторного взрыва оксидазы, которое кодирует ключевую ферментативную субъединицу NADPH-оксидазы растений (Torres and Dangl, 2005). Bvu-miR8 активирует транскрипцию гистоновой ацетилтрансферазы GCN5 в A. thalianaina (Benhamed et al., 2008). Другой белок-мишень MYB6 действует как сигнальный компонент немедленной и положительной активации активного состояния иммунных рецепторов MLA во время перепрограммирования транскрипции для защитных реакций (Chang et al.
Например, экспрессия Bvu-miR1 (белок ARABIDILLO) в A. thaliana регулирует развитие многоклеточных корней (Moody et al., 2012). Bvu-miR2 регулирует экспрессию АТФазы во время развития растений и координирует ее индукцию в ответ на сильное засоление (Lehr et al., 1999). Посредством регуляции транскрипции он также влияет на АТФазную активность субъединицы I хелатазы магния в ячмене (Lake et al., 2004) и на нечувствительность к абсцизовой кислоте 5, необходимую для задержки роста проросших проростков в условиях стресса окружающей среды (Liu and Stone, 2013). Bvu-miR7 нацелен на семейство генов респираторного взрыва оксидазы, которое кодирует ключевую ферментативную субъединицу NADPH-оксидазы растений (Torres and Dangl, 2005). Bvu-miR8 активирует транскрипцию гистоновой ацетилтрансферазы GCN5 в A. thalianaina (Benhamed et al., 2008). Другой белок-мишень MYB6 действует как сигнальный компонент немедленной и положительной активации активного состояния иммунных рецепторов MLA во время перепрограммирования транскрипции для защитных реакций (Chang et al. , 2013). Белки, связывающие элементы, реагирующие на дегидратацию, образуют основное семейство белков, связывающих элементы, чувствительные к AP2/этилену, и играют решающую роль в регуляции реакций на абиотический стресс (Sun et al., 2008). Bvu-miR13 нацеливается на белки WD-повтора в этом разнообразном семействе регуляторных белков. На сегодняшний день полногеномная характеристика этого семейства была проведена только для Arabidopsis , и мало что известно о генах, кодирующих белок WD-повтора, у других видов. Недавно стало известно, что белок WD-repeat играет важную роль в стрессоустойчивости огурцов (Li et al., 2014). Другие мишени, например, богатые лейцином белки повторов, играют важную роль в сигнальных путях как животных, так и растений, регулирующих рост, развитие, дифференцировку, гибель клеток и патогенные защитные реакции (Gou et al., 2010). Эти исследования дали представление о молекулярных механизмах микроРНК и могут иметь большой потенциал для улучшения сахарной свеклы.
, 2013). Белки, связывающие элементы, реагирующие на дегидратацию, образуют основное семейство белков, связывающих элементы, чувствительные к AP2/этилену, и играют решающую роль в регуляции реакций на абиотический стресс (Sun et al., 2008). Bvu-miR13 нацеливается на белки WD-повтора в этом разнообразном семействе регуляторных белков. На сегодняшний день полногеномная характеристика этого семейства была проведена только для Arabidopsis , и мало что известно о генах, кодирующих белок WD-повтора, у других видов. Недавно стало известно, что белок WD-repeat играет важную роль в стрессоустойчивости огурцов (Li et al., 2014). Другие мишени, например, богатые лейцином белки повторов, играют важную роль в сигнальных путях как животных, так и растений, регулирующих рост, развитие, дифференцировку, гибель клеток и патогенные защитные реакции (Gou et al., 2010). Эти исследования дали представление о молекулярных механизмах микроРНК и могут иметь большой потенциал для улучшения сахарной свеклы. Функции этих интересных микроРНК в сахарной свекле необходимо исследовать в будущем.
Функции этих интересных микроРНК в сахарной свекле необходимо исследовать в будущем.
Картирование QTL устойчивости сахарной свеклы к болезням
Пятнистость листьев является одним из наиболее серьезных и широко распространенных лиственных заболеваний сахарной свеклы.Он вызывает некротические поражения и прогрессирующее разрушение структуры и функции листьев растения (Holtschulte, 2000). Болезнь сильно повлияла на урожайность и сахаристость урожая. Удвоенные гаплоиды (DH), популяции F2 рекомбинантных инбредных линий (RILS) и почти изогенные линии (NILS) являются подходящими популяциями для картирования локусов количественных признаков (QTL) (Ibrahim et al., 2012). Чтобы справиться со сложным наследованием устойчивости к пятнистости листьев Cercospora (CLS) , Taguchi et al.(2011) использовали RIL сахарной свеклы, полученные в результате скрещивания устойчивой линии («NK-310mm-O») и восприимчивой («NK-184mm-O») линии. Затем эти RIL были протестированы на устойчивость к патогену CLS в полевых условиях (Taguchi et al. , 2011). Составное картирование интервалов (CIM) показало четыре QTL, вовлеченных в устойчивость к CLS, которые были последовательно обнаружены. Было два устойчивых QTL (qcr1 на хромосоме III, qcr4 на хромосоме IX), которые способствовали устойчивости при скрещивании линий («NK-310mm-O»).Имелись еще два QTL (qcr2 на хромосоме IV, qcr3 на хромосоме VI), которые способствовали развитию устойчивости восприимчивой линии. Кроме того, на хромосоме III в геноме сахарной свеклы картирован ряд важных кластеров генов устойчивости, например: QTL устойчивости к CLS (Setiawan et al., 2000), гены Rz1–Rz5 (Grimmer et al., 2007). ), ген Acr1 (Taguchi et al., 2010), ген RGAs (Lein et al., 2007) и ген X, восстановитель фертильности для CMS Owen (Hagihara et al., 2005). Эти кластеры генов устойчивости на хромосоме III в основном ответственны за устойчивость сахарной свеклы к болезням.
, 2011). Составное картирование интервалов (CIM) показало четыре QTL, вовлеченных в устойчивость к CLS, которые были последовательно обнаружены. Было два устойчивых QTL (qcr1 на хромосоме III, qcr4 на хромосоме IX), которые способствовали устойчивости при скрещивании линий («NK-310mm-O»).Имелись еще два QTL (qcr2 на хромосоме IV, qcr3 на хромосоме VI), которые способствовали развитию устойчивости восприимчивой линии. Кроме того, на хромосоме III в геноме сахарной свеклы картирован ряд важных кластеров генов устойчивости, например: QTL устойчивости к CLS (Setiawan et al., 2000), гены Rz1–Rz5 (Grimmer et al., 2007). ), ген Acr1 (Taguchi et al., 2010), ген RGAs (Lein et al., 2007) и ген X, восстановитель фертильности для CMS Owen (Hagihara et al., 2005). Эти кластеры генов устойчивости на хромосоме III в основном ответственны за устойчивость сахарной свеклы к болезням.
ВАС-библиотека из геномной ДНК хромосомы № 9 сахарной свеклы М14
Компетентная к трансформации растений бинарная ВАС-библиотека была сконструирована из геномной ДНК хромосомы № 9 сахарной свеклы М14 (Fang et al. , 2004). Всего было получено 2365 положительных клонов, которые были помещены в подбиблиотеку, специфичную для хромосомы 9 B. corolliflora (обозначенной bcBAC-IX). Подбиблиотеку bcBAC-IX дополнительно подвергали скринингу с помощью субтрактивного пула кДНК, полученного из семяпочек M14 и цветочных почек B.vulgaris методом подавляющей субтрактивной гибридизации (SSH). Было получено 103 положительных бинарных ВАС, которые потенциально могут содержать гены чужеродной хромосомы № 9, специфически экспрессирующейся во время развития семязачатка и эмбриона М14, и которые могут быть связаны с апомиктической репродукцией. Бинарные клоны BAC полезны для идентификации генов, ответственных за апомиксы, путем генетической трансформации.
, 2004). Всего было получено 2365 положительных клонов, которые были помещены в подбиблиотеку, специфичную для хромосомы 9 B. corolliflora (обозначенной bcBAC-IX). Подбиблиотеку bcBAC-IX дополнительно подвергали скринингу с помощью субтрактивного пула кДНК, полученного из семяпочек M14 и цветочных почек B.vulgaris методом подавляющей субтрактивной гибридизации (SSH). Было получено 103 положительных бинарных ВАС, которые потенциально могут содержать гены чужеродной хромосомы № 9, специфически экспрессирующейся во время развития семязачатка и эмбриона М14, и которые могут быть связаны с апомиктической репродукцией. Бинарные клоны BAC полезны для идентификации генов, ответственных за апомиксы, путем генетической трансформации.
Транскриптомные исследования сахарной свеклы
Применение подавления субтрактивной гибридизации (SSH) в сахарной свекле
SSH — это метод, используемый для идентификации дифференциально экспрессируемых генов в клетках, важных для роста и дифференцировки (Lukyanov et al. , 2007). Этот метод часто применялся для изучения молекулярных механизмов растений при биотических и абиотических стрессах (Sahebi et al., 2015). Реакция на насекомых-вредителей в корнях сахарной свеклы является интересной областью исследований в области защиты растений.
, 2007). Этот метод часто применялся для изучения молекулярных механизмов растений при биотических и абиотических стрессах (Sahebi et al., 2015). Реакция на насекомых-вредителей в корнях сахарной свеклы является интересной областью исследований в области защиты растений.
Puthoff и соавторы идентифицировали более 150 EST корней сахарной свеклы, обогащенных генами, которые реагируют на кормление личинками корней сахарной свеклы как в генотипе F1016 с умеренной устойчивостью, так и в генотипе F1010 с умеренной устойчивостью с использованием SSH [49]. Дифференциальная экспрессия корневых EST была подтверждена с помощью RT-PCR.EST были дополнительно охарактеризованы с использованием профилей экспрессии, созданных на микрочипах, в корнях сахарной свеклы F1016 после механического повреждения и обработки сигнальными молекулами метилжасмонатом, салициловой кислотой и этиленом. Из исследованных ЭСТ 20% регулировались метилжасмонатом, 17% — салициловой кислотой и 11% — этиленом, что позволяет предположить, что эти сигнальные пути участвуют в защитной реакции корней сахарной свеклы. Идентификация этих EST корней сахарной свеклы дает знания о защите корней растений и, вероятно, приведет к разработке новых стратегий борьбы с корневой личинкой сахарной свеклы (Puthoff and Smigocki, 2007).
Идентификация этих EST корней сахарной свеклы дает знания о защите корней растений и, вероятно, приведет к разработке новых стратегий борьбы с корневой личинкой сахарной свеклы (Puthoff and Smigocki, 2007).
SSH также применяли для выделения экспрессируемых генов стержневого корня из сахарной свеклы (Kloos et al., 2002). Стержневой корень сахарной свеклы ( Beta vulgaris L.) претерпевает специфический переход в развитии, чтобы функционировать как запасающий орган. SSH использовали для выделения фрагментов кДНК генов, экспрессируемых в стержневом корне. Молекулярный анализ шести кДНК, кодирующих полные генные продукты, показал, что эти гены содержат гомологи индуцируемого засухой линкерного гистона, основного латексоподобного белка, фосфоенолпируваткарбоксилазной киназы, предполагаемого фермента процессинга вакуолей, тауматин-подобного белка и аланинового белка. — и белок, богатый глутаминовой кислотой.Все эти гены транскрибируются в стержневых корнях, в то время как их экспрессия в листьях низкая или не обнаруживается. SSH также использовался в сахарной свекле M14 для идентификации дифференциально экспрессируемых генов. Субтрактивную библиотеку кДНК получали с помощью SSH между органами цветка M14 и B. vulgaris (Ma et al., 2011). Всего в библиотеке было идентифицировано 190 уникальных последовательностей, и их предполагаемые функции были проанализированы с помощью Gene Ontology (GO). Все EST предоставляют информацию о генах-кандидатах, полезных для изучения репродуктивного развития M14.Один из генов, обозначенный как BvM14-MADS box , кодирует транскрипционный фактор MADS box. Он был клонирован из М14 и сверхэкспрессирован в трансгенных растениях табака. Сверхэкспрессия BvM14-MADS box приводила к значительным фенотипическим изменениям табака (Ma et al., 2011).
SSH также использовался в сахарной свекле M14 для идентификации дифференциально экспрессируемых генов. Субтрактивную библиотеку кДНК получали с помощью SSH между органами цветка M14 и B. vulgaris (Ma et al., 2011). Всего в библиотеке было идентифицировано 190 уникальных последовательностей, и их предполагаемые функции были проанализированы с помощью Gene Ontology (GO). Все EST предоставляют информацию о генах-кандидатах, полезных для изучения репродуктивного развития M14.Один из генов, обозначенный как BvM14-MADS box , кодирует транскрипционный фактор MADS box. Он был клонирован из М14 и сверхэкспрессирован в трансгенных растениях табака. Сверхэкспрессия BvM14-MADS box приводила к значительным фенотипическим изменениям табака (Ma et al., 2011).
Ли и др. (2009) сообщили о сравнительном протеомном и транскриптомном исследовании половых и апомиктических процессов у сахарной свеклы. Библиотеки кДНК были сконструированы с использованием SSH с апомиктической моносомной линией добавления M14 в качестве тестера и B. vulgaris в качестве драйвера. Сравнительный анализ протеомных данных и транскриптомных данных показал, что восемь белков имеют значительное соответствие между уровнями экспрессии белка и мРНК. Большинство совпадающих белков были связаны с метаболизмом. Интересно, что два из соответствующих белков, цистатин и тиоредоксинпероксидаза, были связаны с болезнью и защитной реакцией, что указывает на то, что белки, связанные с защитой, могут участвовать в апомиктическом репродуктивном процессе. Ян и др.(2012) сообщили о транскриптомном анализе листьев и корней сахарной свеклы М14, которые обрабатывались 500 мМ NaCl в течение 7 дней. Технология SSH была использована для получения высококачественной субтрактивной библиотеки кДНК. В общей сложности 600 положительных клонов были случайным образом отобраны и подвергнуты секвенированию ДНК, и было получено 499 неизбыточных EST. После сборки было получено 58 унигенов, в том числе 14 синглетонов и 44 контига. Некоторые чувствительные к соли гены были идентифицированы как важные для метаболизма (например, саденозилметионинсинтаза 2 (SAMS2) и нитритредуктаза), фотосинтеза (например,g.
vulgaris в качестве драйвера. Сравнительный анализ протеомных данных и транскриптомных данных показал, что восемь белков имеют значительное соответствие между уровнями экспрессии белка и мРНК. Большинство совпадающих белков были связаны с метаболизмом. Интересно, что два из соответствующих белков, цистатин и тиоредоксинпероксидаза, были связаны с болезнью и защитной реакцией, что указывает на то, что белки, связанные с защитой, могут участвовать в апомиктическом репродуктивном процессе. Ян и др.(2012) сообщили о транскриптомном анализе листьев и корней сахарной свеклы М14, которые обрабатывались 500 мМ NaCl в течение 7 дней. Технология SSH была использована для получения высококачественной субтрактивной библиотеки кДНК. В общей сложности 600 положительных клонов были случайным образом отобраны и подвергнуты секвенированию ДНК, и было получено 499 неизбыточных EST. После сборки было получено 58 унигенов, в том числе 14 синглетонов и 44 контига. Некоторые чувствительные к соли гены были идентифицированы как важные для метаболизма (например, саденозилметионинсинтаза 2 (SAMS2) и нитритредуктаза), фотосинтеза (например,g. , белок 8, связывающий хлоропластный хлорофилл a–b), энергию (например, фосфоглицераткиназу), синтез белка (например, 60S рибосомный белок L19-3) и расщепление (например, цистеиновая протеаза и протеаза процессинга карбоксильного конца), стресс и защита [например, глутатион-S-трансфераза (GST)]. Это исследование выявило гены-кандидаты для подробной функциональной характеристики и заложило основу для дальнейшего изучения механизмов солеустойчивости у сахарной свеклы.
, белок 8, связывающий хлоропластный хлорофилл a–b), энергию (например, фосфоглицераткиназу), синтез белка (например, 60S рибосомный белок L19-3) и расщепление (например, цистеиновая протеаза и протеаза процессинга карбоксильного конца), стресс и защита [например, глутатион-S-трансфераза (GST)]. Это исследование выявило гены-кандидаты для подробной функциональной характеристики и заложило основу для дальнейшего изучения механизмов солеустойчивости у сахарной свеклы.
Транскриптомика сахарной свеклы в ответ на низкотемпературный стресс
Сравнительная транскриптомика используется для выявления различий в количестве транскриптов между различными сортами, органами, стадиями развития и/или условиями обработки (Mardis, 2008; Schuster, 2008; Bräutigam and Gowik, 2010).Низкотемпературный стресс является существенным фактором, влияющим на качество урожая и вызывающим потери продукции в сельском хозяйстве. Выживаемость молодых всходов сахарной свеклы и последующая урожайность взрослых растений часто серьезно ограничиваются низкой температурой, особенно когда растения подвергаются воздействию отрицательных температур на ранних стадиях развития. Молитерни и др. (2015) определили транскриптомные изменения с помощью высокопроизводительного секвенирования РНК листьев и корней (RNA-Seq) сахарной свеклы, подвергшейся холодовому стрессу, который имитировал условия весенних ночей, иногда переживаемые молодыми саженцами (Moliterni et al., 2015). В ткани корня CBF3 активируется в течение нескольких минут холодового стресса. Авторы предположили, что транскрипция CBF3 в стрессовых растениях либо сохраняется в течение более длительного периода, либо начинается раньше в корнях по сравнению с листьями. Также было обнаружено, что гены семейства AP2/ERF либо активируются, либо активируются во всех органах при холодовом стрессе. Это ожидаемый результат, так как известно, что эти ТФ быстро индуцируются при воздействии низкой температуры на Arabidopsis (Lee et al., 2005). AP2/ERF TFs участвуют в регуляции первичного и специализированного метаболизма и в ряде ответов JA (Licausi et al., 2013).
Молитерни и др. (2015) определили транскриптомные изменения с помощью высокопроизводительного секвенирования РНК листьев и корней (RNA-Seq) сахарной свеклы, подвергшейся холодовому стрессу, который имитировал условия весенних ночей, иногда переживаемые молодыми саженцами (Moliterni et al., 2015). В ткани корня CBF3 активируется в течение нескольких минут холодового стресса. Авторы предположили, что транскрипция CBF3 в стрессовых растениях либо сохраняется в течение более длительного периода, либо начинается раньше в корнях по сравнению с листьями. Также было обнаружено, что гены семейства AP2/ERF либо активируются, либо активируются во всех органах при холодовом стрессе. Это ожидаемый результат, так как известно, что эти ТФ быстро индуцируются при воздействии низкой температуры на Arabidopsis (Lee et al., 2005). AP2/ERF TFs участвуют в регуляции первичного и специализированного метаболизма и в ряде ответов JA (Licausi et al., 2013). Сообщалось, что недостаток ADA2b TF приводит к увеличению устойчивости к замораживанию, влияя на занятость нуклеосом у Arabidopsis (Vlachonasios et al., 2003). Кроме того, предполагаемая гистоновая ацетилаза и лизин-специфическая деметилаза сильно активируются в листьях при холодовом стрессе, что указывает на ремоделирование хроматина и модификацию ответа.
Сообщалось, что недостаток ADA2b TF приводит к увеличению устойчивости к замораживанию, влияя на занятость нуклеосом у Arabidopsis (Vlachonasios et al., 2003). Кроме того, предполагаемая гистоновая ацетилаза и лизин-специфическая деметилаза сильно активируются в листьях при холодовом стрессе, что указывает на ремоделирование хроматина и модификацию ответа.
Эти исследования показали, что метаболический путь, наиболее подверженный влиянию низкой температуры, связан с углеводным обменом. Кроме того, авторы обнаружили 13 дифференциально экспрессируемых последовательностей, связанных со вторичным метаболизмом фосфолипидов, ни одна из которых не была общей для листьев и корней, что указывает на то, что эти пути являются еще одним важным компонентом ранней передачи холодовых сигналов в корнях сахарной свеклы. Высокая степень органоспецифичности, вероятно, связана с репертуаром соединений, синтезируемых двумя органами при стрессе.Эти данные пролили свет на транскриптом молодой сахарной свеклы во время холодового стресса в ночное время и подробно описали как органоспецифичность, так и общие пути физиологической реакции на низкие температуры. Эти методы транскриптомики, основанные на RNA-Seq, являются эффективным и мощным инструментом, с анализом, выявляющим новые гены для будущих исследований.
Эти методы транскриптомики, основанные на RNA-Seq, являются эффективным и мощным инструментом, с анализом, выявляющим новые гены для будущих исследований.
Протеомные исследования сахарной свеклы
Протеомный анализ был проведен для решения нескольких важных вопросов во многих процессах, таких как: передача сигналов, регуляторные процессы и транспорт в растениях (Zhang et al., 2008). Важные сведения о протеомной реакции на стресс были в основном получены в результате исследований модельных растений A. thaliana и риса (Janmohammadi et al., 2015; Liu et al., 2015; Xu et al., 2015). Протеомный анализ обеспечивает важный способ проверки изменений уровня белка, чтобы помочь идентифицировать новые белки.
Белки различных сортов сахарной свеклы без обработки стрессом
Zhu et al. (2009) сравнили протеомы моносомной добавочной линии М14 и В.vulgaris с использованием 2-DE (двумерный гель-электрофорез). Они идентифицировали 27 белковых пятен с помощью MALDI-TOF MS.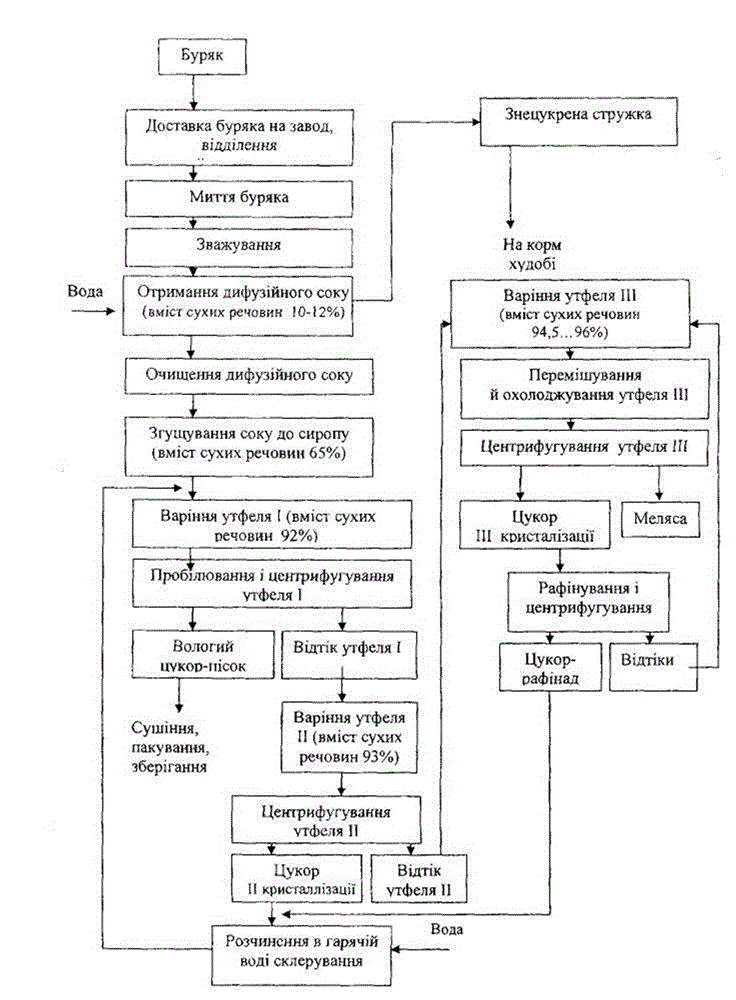 Среди них только два белковых пятна были обнаружены у B. vulgaris и пять белковых пятен были уникальными для М14. Эти белки были вовлечены во многие биологические пути. Результаты могут быть полезны для нас, чтобы лучше понять, как различия генотипов связаны с различиями протеома и фенотипа.
Среди них только два белковых пятна были обнаружены у B. vulgaris и пять белковых пятен были уникальными для М14. Эти белки были вовлечены во многие биологические пути. Результаты могут быть полезны для нас, чтобы лучше понять, как различия генотипов связаны с различиями протеома и фенотипа.
Ли и др. (2009) представили сравнительный протеомный и транскриптомный анализ половых и апомиктических процессов у сахарной свеклы.Всего в ходе апомиктического репродуктивного развития с помощью 2-ДЭ и МС-анализа было идентифицировано 71 дифференциально экспрессируемое белковое пятно из цветочных органов М14. Дифференциально экспрессируемые белки были вовлечены в несколько процессов, которые могут работать совместно, чтобы способствовать апомиктической репродукции, создавая новые потенциальные белковые маркеры, важные для апомиктического развития.
Изменения протеома в ответ на солевой стресс у сахарной свеклы
На сегодняшний день имеются сообщения о некоторых протеомных исследованиях, касающихся реакции сахарной свеклы на солевой стресс. Уэйкил и др. идентифицировали шесть белков из побегов сахарной свеклы и три белка из корней, которые значительно изменились при обработке 125 мМ соли (Wakeel et al., 2011). Наша группа провела протеомный анализ моносомной аддитивной линии М14 в условиях солевого стресса 500 мМ в течение 7 дней. В общей сложности с помощью ЖХ-МС/МС было идентифицировано 71 дифференциально экспрессируемое белковое пятно 2D. Самая большая функциональная группа представлена метаболизмом (28%), за ней следуют энергия (21%), синтез белка (10%), стресс и защита (10%), белки назначения (8%), неизвестные белки (8%), вторичный метаболизм (5%), сигнальная трансдукция (4%), транспортеры (1%) и деление клеток (1%).Из идентифицированных белков только восемь имели соответствующие транскриптомные данные. Это подчеркивает важность профилирования экспрессии на белковом уровне (Li et al., 2009). На этом основании мы сосредоточились на функциях цистатина (Wang et al., 2012), глиоксалазы I (Wu et al., 2013), CCoAOMT и тиоредоксинпероксидазы.
Уэйкил и др. идентифицировали шесть белков из побегов сахарной свеклы и три белка из корней, которые значительно изменились при обработке 125 мМ соли (Wakeel et al., 2011). Наша группа провела протеомный анализ моносомной аддитивной линии М14 в условиях солевого стресса 500 мМ в течение 7 дней. В общей сложности с помощью ЖХ-МС/МС было идентифицировано 71 дифференциально экспрессируемое белковое пятно 2D. Самая большая функциональная группа представлена метаболизмом (28%), за ней следуют энергия (21%), синтез белка (10%), стресс и защита (10%), белки назначения (8%), неизвестные белки (8%), вторичный метаболизм (5%), сигнальная трансдукция (4%), транспортеры (1%) и деление клеток (1%).Из идентифицированных белков только восемь имели соответствующие транскриптомные данные. Это подчеркивает важность профилирования экспрессии на белковом уровне (Li et al., 2009). На этом основании мы сосредоточились на функциях цистатина (Wang et al., 2012), глиоксалазы I (Wu et al., 2013), CCoAOMT и тиоредоксинпероксидазы.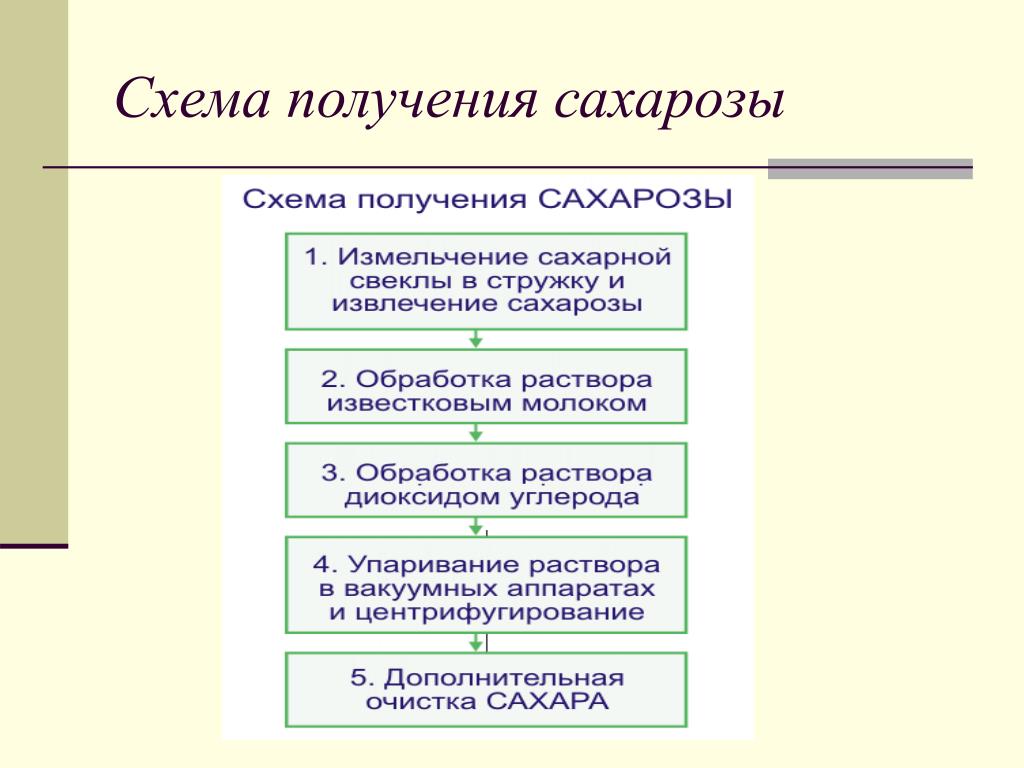 Все эти белки показали повышенный уровень белка при солевом стрессе. Трансгенные растения проявляли повышенную устойчивость к солевому стрессу. Это исследование напрямую улучшило наше понимание механизмов, лежащих в основе высокой солеустойчивости M14.
Все эти белки показали повышенный уровень белка при солевом стрессе. Трансгенные растения проявляли повышенную устойчивость к солевому стрессу. Это исследование напрямую улучшило наше понимание механизмов, лежащих в основе высокой солеустойчивости M14.
Другое протеомное исследование направлено на выявление солечувствительных белков в растениях M14 в условиях мягкого солевого стресса с концентрацией 0, 200 и 400 мМ NaCl с использованием 2D-DIGE для отделения белков от контрольных и обработанных солью листьев и корней M14 (Yang et др., 2013). Дифференциально экспрессируемые белки идентифицировали с помощью нанопоточной жидкостной хроматографии (ЖХ)-МС/МС и поиска в базе данных Mascot. В качестве дополнительного подхода использовали iTRAQ LC-MS/MS для идентификации и количественного определения дифференциально экспрессируемых белков во время реакции на соленость в M14.Мы идентифицировали 86 белковых пятен, представляющих 67 уникальных белков в листьях, и 22 белковых пятна, представляющих 22 уникальных белка в корнях. Кроме того, в листьях было идентифицировано 75 дифференциально экспрессируемых белков, а в корнях – 43 дифференциально экспрессируемых белка соответственно. Белки в основном участвовали в фотосинтезе, энергии, метаболизме, сворачивании и деградации белков, а также в стрессе и защите. По сравнению с транскриптомными данными, 13 белков в листьях и 12 белков в корнях показали значительную корреляцию в экспрессии генов и уровнях белка.Эти результаты позволяют предположить, что существует несколько процессов, лежащих в основе устойчивости М14 к солевому стрессу.
Кроме того, в листьях было идентифицировано 75 дифференциально экспрессируемых белков, а в корнях – 43 дифференциально экспрессируемых белка соответственно. Белки в основном участвовали в фотосинтезе, энергии, метаболизме, сворачивании и деградации белков, а также в стрессе и защите. По сравнению с транскриптомными данными, 13 белков в листьях и 12 белков в корнях показали значительную корреляцию в экспрессии генов и уровнях белка.Эти результаты позволяют предположить, что существует несколько процессов, лежащих в основе устойчивости М14 к солевому стрессу.
Наша группа также сообщила об изменениях протеома мембран растений М14 в ответ на солевой стресс (0, 200, 400 мМ NaCl; Li H. et al., 2015). Мы использовали двумерную технологию iTRAQ LC-MS/MS для количественного протеомного анализа. Всего было идентифицировано 274 белка, и большинство из них были мембранными белками. Всего было идентифицировано 50 дифференциальных белков, из которых 40 белков демонстрировали повышенную экспрессию, а 10 — пониженную экспрессию.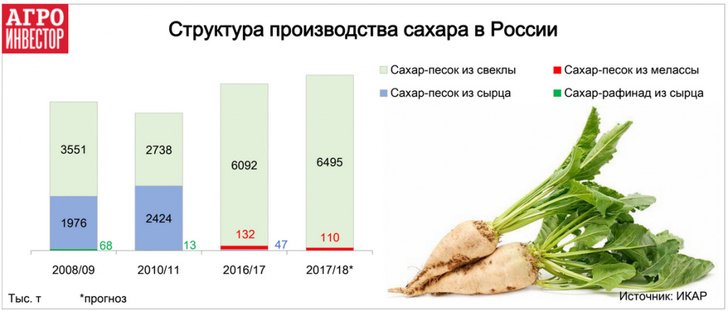 Белки в основном участвовали в транспорте (17%), метаболизме (16%), синтезе белков (15%), фотосинтезе (13%), фолдинге и деградации белков (9%), передаче сигналов (6%), стрессе и защите. (6%), энергия (6%) и клеточная структура (2%). Эти результаты показали, что мембранные белки вносят вклад в толерантность к солевому стрессу, наблюдаемую у М14.
Белки в основном участвовали в транспорте (17%), метаболизме (16%), синтезе белков (15%), фотосинтезе (13%), фолдинге и деградации белков (9%), передаче сигналов (6%), стрессе и защите. (6%), энергия (6%) и клеточная структура (2%). Эти результаты показали, что мембранные белки вносят вклад в толерантность к солевому стрессу, наблюдаемую у М14.
Изменения протеома сахарной свеклы в ответ на стресс от засухи
Hajheidari et al. (2005) изучали изменения протеома сахарной свеклы в ответ на стресс от засухи.Собирали листья с хорошо политых и обработанных засухой растений через 157 дней после посева. Изменения белков анализировали с помощью 2D-DIGE с последующим анализом изображений. Выявлено более 500 белковых пятен, 79 пятен претерпели значительные изменения при засушливом стрессе. Двадцать белковых пятен расщепляли и подвергали ЖХ-МС/МС, и идентифицировали 11 белков, участвующих в окислительном стрессе, передаче сигналов и окислительно-восстановительной регуляции. Эти белки могут быть важными мишенями для улучшения устойчивости растений к абиотическому стрессу посредством селекции.
Метаболомические исследования сахарной свеклы
Метаболомика — это захватывающая технология, которая использовалась для выявления вторичных метаболитов, важных для физиологических процессов и различных реакций на стресс (Capuano et al., 2013). В процессе развития растений и взаимодействия с окружающей средой динамический метаболом отражает физиологические и биохимические процессы растения и может определять фенотипы и признаки (Fernie et al., 2004; Oksman-Caldentay and Saito, 2005). В настоящее время основными аналитическими инструментами в метаболомике являются газовая хроматография (ГХ)–МС, ЖХ–МС, капиллярный электрофорез (КЭ)–МС и ядерно-магнитный резонанс (ЯМР).Казимерчак и др. (2014) определили уровни метаболитов как в сыром корне свеклы, так и в естественно ферментированном свекольном соке из органических (ORG) продуктов по сравнению с обычными (CONV) продуктами. Цель работы состояла в том, чтобы выяснить ценность ферментированного свекольного сока с точки зрения противораковых свойств. Результаты показали, что свежая свекла ORG содержала больше полезных соединений, чем свекла CONV, таких как сухое вещество и витамин С, больше, чем свекла CONV. По сравнению с соком CONV было обнаружено, что ферментированные соки ORG обладают более сильной противораковой активностью.Метаболомика все еще находится в зачаточном состоянии, и такие анализы сахарной свеклы проводятся редко, но можно ожидать, что в будущих исследованиях будет реализована эта мощная технология.
Результаты показали, что свежая свекла ORG содержала больше полезных соединений, чем свекла CONV, таких как сухое вещество и витамин С, больше, чем свекла CONV. По сравнению с соком CONV было обнаружено, что ферментированные соки ORG обладают более сильной противораковой активностью.Метаболомика все еще находится в зачаточном состоянии, и такие анализы сахарной свеклы проводятся редко, но можно ожидать, что в будущих исследованиях будет реализована эта мощная технология.
Важные области применения сахарной свеклы будут улучшены с помощью OMICS
Многие растения накапливают глицин-бетаин (бетаин) для регулирования биохимических и физиологических процессов в условиях абиотических стрессов (Takabe et al., 2006). Например, глицин-бетаин служит донором метила в нескольких биохимических путях (Pummer et al., 2000).Сахарная свекла представляет собой накапливающее бетаин двудольное растение с высокой экономической ценностью (Catusse et al., 2008). Сообщалось, что бетаин синтезируется путем двухстадийного окисления холина, при котором холинмонооксигеназа (ХМО) катализирует первую стадию, а бетаинальдегиддегидрогеназа (БАДГ) выполняет вторую стадию (Yamada et al. , 2009). Таким образом, CMO является ключевым ферментом для защиты растений от абиотических стрессов. Он был обнаружен у Chenopodiaceae и Amaranthaceae, но не у некоторых растений, накапливающих бетаин, таких как мангровые заросли (Bhuiyan et al., 2007). В отличие от сахарной свеклы, у многих растений нет пути биосинтеза бетаина. Таким образом, генная инженерия путей биосинтеза бетаина представляет собой потенциальный способ улучшить стрессоустойчивость растений (Hibino et al., 2001; Fitzgerald et al., 2009).
, 2009). Таким образом, CMO является ключевым ферментом для защиты растений от абиотических стрессов. Он был обнаружен у Chenopodiaceae и Amaranthaceae, но не у некоторых растений, накапливающих бетаин, таких как мангровые заросли (Bhuiyan et al., 2007). В отличие от сахарной свеклы, у многих растений нет пути биосинтеза бетаина. Таким образом, генная инженерия путей биосинтеза бетаина представляет собой потенциальный способ улучшить стрессоустойчивость растений (Hibino et al., 2001; Fitzgerald et al., 2009).
Помимо бетаина, беталаины богаты красной свеклой и существуют только в 10 семействах растений Caryophyllale . Красная свекла ( B. vulgaris ) широко используется в качестве пищевого ингредиента из-за ее свекольно-красного цвета.Поэтому большинство исследований компонентов красной свеклы были сосредоточены на беталаинах. В настоящее время выход беталаинов, извлеченных и очищенных из красной свеклы, составляет всего около 10%. Было показано, что фенольные смолы (такие как беталаины) обладают питательной ценностью, и растет интерес к использованию этих растительных компонентов для улучшения пищевых ингредиентов и в качестве антиоксидантов. Беталаины являются важными растительными фенолами со многими привлекательными свойствами, такими как: стабильность, антиоксидантная активность, противоопухолевые свойства и снижение уровня липидов и сахара в крови.Кроме того, беталаины являются эффективными поглотителями свободных радикалов, которые помогают поддерживать здоровье и защищать от таких заболеваний, как рак и ишемическая болезнь сердца (Kujala et al., 2000; Han et al., 2015; Mikołajczyk-Bator et al., 2016). ).
Беталаины являются важными растительными фенолами со многими привлекательными свойствами, такими как: стабильность, антиоксидантная активность, противоопухолевые свойства и снижение уровня липидов и сахара в крови.Кроме того, беталаины являются эффективными поглотителями свободных радикалов, которые помогают поддерживать здоровье и защищать от таких заболеваний, как рак и ишемическая болезнь сердца (Kujala et al., 2000; Han et al., 2015; Mikołajczyk-Bator et al., 2016). ).
Кроме того, сахарная свекла обеспечивает примерно 30% годового производства сахара в мире и является источником как биоэтанола, так и корма для животных. Дхар и др. (2015) разработали два высокоэффективных метода производства газообразного водорода из сока сахарной свеклы в качестве источника чистой энергии (Dhar et al., 2015). Побочные продукты сахарной свеклы (ППС), образующиеся при промышленном извлечении сахара, в основном состоят из жома и мелассы, и использование ППБ в качестве возобновляемого источника энергии может обеспечить дополнительные экономические и экологические преимущества (Aboudi et al. , 2015).
, 2015).
Исследования OMICS могут значительно улучшить потенциальное применение сахарной свеклы, по крайней мере, тремя способами. Одним из них является улучшение наших знаний о молекулярных сетях, связанных с синтезом ключевых метаболитов, например, бетаина глицина и беталаинов. Эти знания позволят моделировать и обосновывать разработку важных метаболитов.Кроме того, мы можем использовать OMICS для изучения глобальных молекулярных изменений, происходящих в ответ на стресс, и устойчивости сахарной свеклы к стрессовым условиям. Эта информация может помочь улучшить стрессоустойчивость и тем самым повысить урожайность сахарной свеклы даже в неидеальных условиях. Наконец, исследования уникальных зародышевых плазм сахарной свеклы (например, M14 в условиях солевого стресса) могут быть полезны для повышения урожайности и производства продуктов питания и биоэнергии другими культурами.
Заключение
В этом обзоре мы обобщили технологии OMICS и их применение в сахарной свекле, включая: M14 для идентификации новых генов, белков, связанных с биотическими и абиотическими стрессами, апомиксов и метаболитов, связанных с энергией, пищей и здоровьем человека. Геномика — это мощная технология, позволяющая составить план полного генома сахарной свеклы. Механизмы, лежащие в основе апомиксов и стрессоустойчивости, в основном изучались с использованием технологий транскриптомики и протеомики, в то время как исследования метаболомики сахарной свеклы все еще редки. К настоящему времени идентифицировано множество генов и белков, связанных с апомиксами и солевым стрессом, для выявления апомиксов и механизмов солеустойчивости у специальной гермоплазмы сахарной свеклы М14. Результаты расширили наше понимание молекулярных механизмов реакции сахарной свеклы на устойчивость к биотическим и абиотическим стрессам и апомиксам, которые могут быть применены для повышения стрессоустойчивости сахарной свеклы и других культур для улучшения производства продуктов питания, производства энергии (например,например, газообразный водород и биоэтанол), а также накопление полезных для здоровья химических веществ (таких как беталаины). Несмотря на то, что использование сахарной свеклы для производства чистой энергии газообразного водорода и биоэтанола, а также для выделения беталаинов, используемых в натуральных пищевых красителях, пищевых добавках и лекарствах, не получило широкого распространения на рынке, сахарная свекла как культура с высокой экономической ценностью будет иметь процветающую перспективу.
Геномика — это мощная технология, позволяющая составить план полного генома сахарной свеклы. Механизмы, лежащие в основе апомиксов и стрессоустойчивости, в основном изучались с использованием технологий транскриптомики и протеомики, в то время как исследования метаболомики сахарной свеклы все еще редки. К настоящему времени идентифицировано множество генов и белков, связанных с апомиксами и солевым стрессом, для выявления апомиксов и механизмов солеустойчивости у специальной гермоплазмы сахарной свеклы М14. Результаты расширили наше понимание молекулярных механизмов реакции сахарной свеклы на устойчивость к биотическим и абиотическим стрессам и апомиксам, которые могут быть применены для повышения стрессоустойчивости сахарной свеклы и других культур для улучшения производства продуктов питания, производства энергии (например,например, газообразный водород и биоэтанол), а также накопление полезных для здоровья химических веществ (таких как беталаины). Несмотря на то, что использование сахарной свеклы для производства чистой энергии газообразного водорода и биоэтанола, а также для выделения беталаинов, используемых в натуральных пищевых красителях, пищевых добавках и лекарствах, не получило широкого распространения на рынке, сахарная свекла как культура с высокой экономической ценностью будет иметь процветающую перспективу. применения в пищевой, биоэнергетической и фармацевтической промышленности.
применения в пищевой, биоэнергетической и фармацевтической промышленности.
Вклад авторов
YZ собрал и проанализировал ссылки для этой статьи и написал первый черновик, JN нарисовал рисунок и помог в организации ссылок.BY играл надзорную роль, руководил написанием и организацией. Все три автора редактировали рукопись.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Это исследование было поддержано Национальным научным фондом Китая (проект 31471552: Реакция антиоксидантных ферментов на солевой стресс в сахарной свекле M14 и проект 31501359: Исследование сайтов фосфорилирования протеинкиназы Ser/Thr из сахара). свеклы M14 в ответ на солевой стресс) и 100 академических программ поддержки позвоночника для молодых преподавателей Университета Хэйлунцзян в 2016 году и Коллектива научно-технических инноваций Общего колледжа провинции Хэйлунцзян (2014TD004).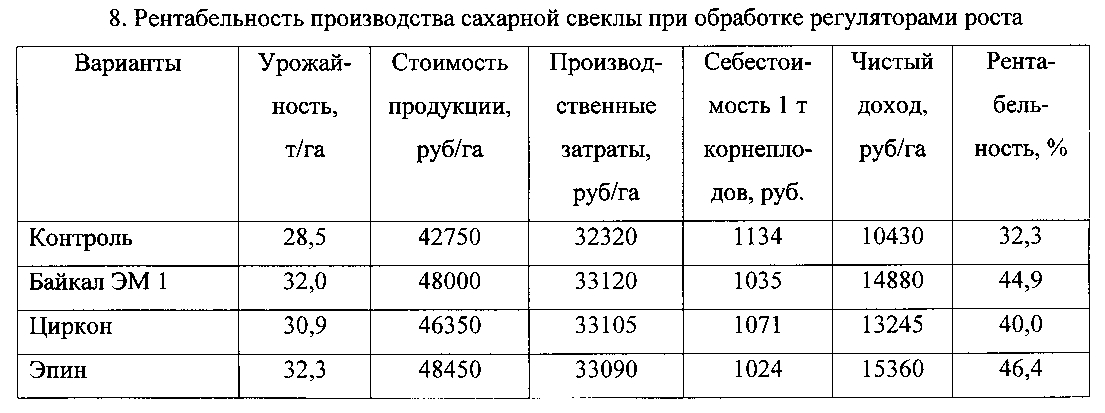 Бумага представляет серийный номер 016 от нашей инновационной группы в Хэйлунцзянском университете (Hdtd2010-05). Мы благодарим Джо Коллинза из отдела молекулярной и клеточной биологии растений Университета Флориды за критическое прочтение рукописи.
Бумага представляет серийный номер 016 от нашей инновационной группы в Хэйлунцзянском университете (Hdtd2010-05). Мы благодарим Джо Коллинза из отдела молекулярной и клеточной биологии растений Университета Флориды за критическое прочтение рукописи.
Ссылки
- Абуди К., Альварес-Гальего С.Х., Ромеро-Гарсия Л.И. (2015). Полунепрерывное анаэробное совместное сбраживание побочных продуктов сахарной свеклы и свиного навоза: влияние скорости органической загрузки (OLR) на производительность процесса. Биоресурс. Технол. 194, 283–290.10.1016/j.biortech.2015.07.031 [PubMed] [CrossRef] [Google Scholar]
- Акпынар Б. А., Кантар М., Будак Х. (2015). Корневые предшественники микроРНК у дикой эммер и современной пшеницы демонстрируют значительные различия в реакции на стресс, вызванный засухой. Функц. интегр. Геномика 15, 587–598. 10.1007/s10142-015-0453-0 [PubMed] [CrossRef] [Google Scholar]
- Барнабас Б., Ягер К., Фехар А. (2008). Влияние засухи и теплового стресса на репродуктивные процессы у злаков. Окружающая среда растительной клетки.
31, 11–38. 10.1111/j.1365-3040.2007.01727.x [PubMed] [CrossRef] [Google Scholar]
- Baum J. A., Bogaert T., Clinton W., Heck G. R., Feldmann P., Ilagan O., et al. . (2007). Борьба с жесткокрылыми насекомыми-вредителями с помощью РНК-интерференции. Нац. Биотехнолог. 25, 1322–1326. 10.1038/nbt1359 [PubMed] [CrossRef] [Google Scholar]
- Benhamed M., Martin-Magniette M.L., Taconnat L., Bitton F., Servet C., De Clercq R., et al. . (2008). Массив промоторов Arabidopsis в масштабе генома идентифицирует мишени гистоновой ацетилтрансферазы GCN5.Плант Дж. 56, 493–504. 10.1111/j.1365-313X.2008.03606.x [PubMed] [CrossRef] [Google Scholar]
- Бенуа И., Чжоу М., Вивас Дуарте А., Даунс Д. Дж., Тодд Р. Б., Клёзен В. и др. . (2015). Пространственная дифференциация экспрессии генов в колониях Aspergillus niger , выращенных для использования свекловичного жома. Научный представитель 5:13592. 10.1038/srep13592 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Бхуйян Н. Х., Хамада А., Ямада Н., Рай В., Хибино Т., Такабэ Т. (2007). Регуляция синтеза бетаина с помощью предшественника и экспрессии холинмонооксигеназы в Amaranthus tricolor.Дж. Эксп. Бот.
58, 4203–4212. 10.1093/jxb/erm278 [PubMed] [CrossRef] [Google Scholar]
- Bräutigam A., Gowik U. (2010). Что секвенирование нового поколения может сделать для вас? Секвенирование следующего поколения как ценный инструмент в исследованиях растений. биол. растений 12, 831–841. 10.1111/j.1438-8677.2010.00373.x [PubMed] [CrossRef] [Google Scholar]
- Brosché M., Vinocur B., Alatalo E.R., Lamminmäki A., Teichmann T., Ottow E.A., et al. . (2005). Экспрессия генов и профилирование метаболитов Populus euphratica , произрастающих в пустыне Негев.Геном биол. 6: Р101. 10.1186/gb-2005-6-12-r101 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Будак Х., Акпынар Б. А. (2015). МикроРНК растений: биогенез, организация и происхождение. Функц. интегр. Геномика
15, 523–531. 10. 1007/s10142-015-0451-2 [PubMed] [CrossRef] [Google Scholar]
- Будак Х., Кантар М., Булут Р., Акпынар Б. А. (2015). Стресс-чувствительные миРНК и изомиР в злаках. Растениевод. 235, 1–13. 10.1016/j.plantsci.2015.02.008 [PubMed] [CrossRef] [Google Scholar]
- Капуано Э., Берригтер-Энлинг Р., ван дер Веер Г., ван Рут С.М. (2013). Аналитическая аутентификация органических продуктов: обзор маркеров. J. Sci. Фуд Агрик. 93, 12–28. 10.1002/jsfa.5914 [PubMed] [CrossRef] [Google Scholar]
- Catusse J., Strub J.M., Job C., Dorsselaer A., Job D. (2008). Протеомная характеристика силы семян сахарной свеклы и ее тканеспецифического выражения. проц. Натл. акад. науч. США. 105, 10262–10267. 10.1073/pnas.0800585105 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Chang C., Ю Д., Цзяо Дж., Цзин С., Шульце-Леферт П., Шен К.Х. (2013). Иммунные рецепторы MLA ячменя напрямую взаимодействуют с антагонистически действующими факторами транскрипции, чтобы инициировать передачу сигналов устойчивости к болезням. Растительная клетка
25, 1158–1173. 10.1105/tpc.113.109942 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Chen T., Li Z., Yin X., Hu F., Hu C. (2016). Дискриминация генетически модифицированной сахарной свеклы на основе терагерцовой спектроскопии. Спектрохим. Акта А Мол. биомол. Спектроск. 153, 586–590.10.1016/j.saa.2015.09.028 [PubMed] [CrossRef] [Google Scholar]
- Кураба Дж., Сингх М. Б., Бхалла П. Л. (2014). миРНК в перекрестных помехах между сигнальными путями фитогормонов. Дж. Эксп. Бот. 65, 1425–1438. 10.1093/jxb/eru002 [PubMed] [CrossRef] [Google Scholar]
- Dalke L., Filutowicz A., Pawelska-Koziuska K. (1971). Результаты исследований на гибридах между тетраплоидной однозародышевой сахарной свеклой и Beta corolliftora 2n-36. биул. Инст. Ход. Аклим. росл. 6, 3–7. [Google Scholar]
- Далмэй Т.(2006). Короткие РНК в адаптации к окружающей среде. проц. биол. науч. 273, 1579–1585. 10.1098/rspb.2006.3516 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Дхар Б. Р., Эльбешбиши Э., Хафез Х., Ли Х. С. (2015). Производство водорода из сока сахарной свеклы с использованием интегрированного биоводородного процесса темного брожения и микробного электролизера. Биоресурсная технология.
198, 223–230. 10.1016/j.biortech.2015.08.048 [PubMed] [CrossRef] [Google Scholar]
- Dohm JC, Minoche A.E., Holtgräwe D., Capella-Gutiérrez S., Zakrzewski F., Tafer H., et al. . (2014). Геном недавно одомашненной сахарной свеклы ( Beta vulgaris ). Природа 505, 546–549. 10.1038/nature12817 [PubMed] [CrossRef] [Google Scholar]
- Дрейкотт А. П. (2006). Сахарная свекла. Оксфорд: Блэквелл. [Google Scholar]
- Дунайска-Ордак К., Скорупа-Клапут М., Курник К., Третин А., Тыбурский Ю. (2014). Клонирование и анализ экспрессии гена, кодирующего аскорбатпероксидазу и реагирующего на солевой стресс у свеклы ( Beta vulgaris ).Завод Мол. биол. Респ. 32, 162–175. 10.1007/s11105-013-0636-6 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Эрген Н. З., Будак Х. (2009). Секвенирование более 13 000 экспрессированных меток последовательностей из шести субтрактивных библиотек кДНК диких и современных пшениц после медленного засушливого стресса. Окружающая среда растительной клетки.
32, 220–236. 10.1111/j.1365-3040.2008.01915.x [PubMed] [CrossRef] [Google Scholar]
- Fang X., Gu S., Xu Z., Chen F., Guo D., Zhang H.B., et al. . (2004). Создание бинарной ВАС-библиотеки для апомиктической моносомной добавочной линии Beta corolliflora в сахарной свекле и идентификация клонов, полученных из чужеродной хромосомы.Теор. заявл. Жене. 108, 1420–1425. 10.1007/s00122-003-1566-8 [PubMed] [CrossRef] [Google Scholar]
- Фернандес-Гарсия Н., Эрнандес М., Касадо-вела Дж., Бру Р., Элорца Ф., Хедден П. и др. др. . (2011). Изменения протеома и целевых метаболитов ксилемсапина Brassica oleracea в ответ на солевой стресс. Окружающая среда растительной клетки. 34, 821–836. 10.1111/j.1365-3040.2011.02285.x [PubMed] [CrossRef] [Google Scholar]
- Fernie A. R., Trethewey R. N., Krotzky A. J., Willmitzer L.(2004). Метаболитное профилирование: от диагностики к системной биологии. Нац. Преподобный Мол. Клеточная биол.
5, 763–769. 10.1038/nrm1451 [PubMed] [CrossRef] [Google Scholar]
- Филутович А., Далке Л. (1976). Межвидовые гибриды секции Corollinae Род Beta. Ход. Росл. Аклим. Нас. 20, 1–17. [Google Scholar]
- Фицджеральд Т. Л., Уотерс Д. Л. Э., Генри Р. Дж. (2009). Бетаинальдегиддегидрогеназа растений. биол. растений 11, 119–130. 10.1111/j.1438-8677.2008.00161.x [PubMed] [CrossRef] [Google Scholar]
- Gou X., He K., Yang H., Yuan T., Lin H., Clouse S.D., et al. . (2010). Полногеномное клонирование и анализ последовательности генов протеинкиназы, подобной рецептору, богатому лейцином, в Arabidopsis thaliana . Геномика BMC 11:19. 10.1186/1471-2164-11-19 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Grimmer M.K., Trybush S., Hanley S., Francis S.A., Karp A. Asher, MJC, et al. . (2007). Карта привязанного сцепления для сахарной свеклы на основе маркеров AFLP, SNP и RAPD и картирование QTL нового источника устойчивости к вирусу некроза желтой жилки свеклы.Теор. заявл. Жене.
114, 1151–1160. 10.1007/s00122-007-0507-3 [PubMed] [CrossRef] [Google Scholar]
- Guo D., Wang G., Kang C., Jia S., Liu L. (1994). Исследования по межвидовому скрещиванию сахарной свеклы культурной и Beta corolliflora Zoss. Китайская сахарная свекла 3, 2–7. [Google Scholar]
- Хагихара Э., Итчода Н., Хабу Ю., Иида С., Миками Т., Кубо Т. (2005). Молекулярное картирование гена восстановителя фертильности для цитоплазматической мужской стерильности Оуэна у сахарной свеклы. Теор. заявл.Жене. 111, 250–255. 10.1007/s00122-005-2010-z [PubMed] [CrossRef] [Google Scholar]
- Хаджхейдари М., Абдоллахян-Ногаби М., Аскари Х., Хейдари М., Садегян С.Ю., Обер Э.С. и др. . (2005). Протеомный анализ листьев сахарной свеклы в условиях засушливого стресса. протеомика
5, 950–960. 10.1002/pmic. 200401101 [PubMed] [CrossRef] [Google Scholar]
- Хан Дж., Тан С., Ван Ю., Ян С., Тан Д. (2015). Бетанин снижает накопление и образование поперечных связей коллагена в сердце крысы с высоким содержанием фруктозы путем ингибирования неферментативного гликирования.хим. биол. Взаимодействовать. 5, 37–44. 10.1016/j.cbi.2014.12.032 [PubMed] [CrossRef] [Google Scholar]
- Hibino T., Meng Y.L., Kawamitsu Y., Uehara N., Matsuda N., Tanaka Y., et al. . (2001). Молекулярное клонирование и функциональная характеристика двух видов бетаин-альдегиддегидрогеназы в бетаин-аккумулирующих мангровых деревьях Avicennia marina (Форск). Завод Мол. биол. 45, 353–363. 10.1023/A:1006497113323 [PubMed] [CrossRef] [Google Scholar]
- Holtschulte B. (2000). Cercospora beticola — всемирное распространение и заболеваемость, Cercospora Beticola Sacc.Биология, агрономическое влияние и меры контроля сахарной свеклы, под редакцией Ашера М. Дж. К., Холтшульте Б., Ричарда М. М. , Россо Ф., Штайнрюкен Г., Беккерса Р. (Бельгия: Международный институт исследований свеклы; ), 5–16. [Google Scholar]
- Ибрагим С. Э., Шуберт А., Пиллен К., Леон Дж. (2012). Сравнение QTL признаков засухоустойчивости между двумя продвинутыми популяциями обратного скрещивания яровой пшеницы. Междунар. акад. Дж. 2, 216–227. [Google Scholar]
- Джанмохаммади М., Золла Л., Ринальдуччи С.(2015). Низкотемпературная толерантность растений: изменения на уровне белка. Фитохимия 117, 76–89. 10.1016/j.phytochem.2015.06.003 [PubMed] [CrossRef] [Google Scholar]
- Jia X., Sun C., Zuo Y., Li G., Li G., Ren L., et al. . (2016). Интеграция транскриптомики и метаболомики для характеристики ответа Astragalus membranaceus Bge. вар. mongolicus (Bge) к прогрессирующему стрессу от засухи . Геномика BMC 17:188. 10.1186/s12864-016-2554-0 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Хорхе Т.F., Rodrigues J.A., Caldana C., Schmidt R., van Dongen J. T., Thomas-Oates J. и соавт. . (2015). Метаболомика растений на основе масс-спектрометрии: реакция метаболитов на абиотический стресс. Масс-спектр. Ред. [Epub перед печатью]. 10.1002/mas.21449 [PubMed] [CrossRef] [Google Scholar]
- Kazimierczak R., Hallmann E., Lipowski J., Drela N., Kowalik A., Püssa T., et al. . (2014). Свекла ( Beta vulgaris L.) и естественно ферментированные свекольные соки органического и традиционного производства: метаболомика, уровень антиоксидантов и противораковая активность.J. Sci. Фуд Агрик. 94, 2618–2629. 10.1002/jsfa.6722 [PubMed] [CrossRef] [Google Scholar]
- Kloos D. U., Oltmanns H., Dock C., Stahl D., Hehl R. (2002). Выделение и молекулярный анализ шести экспрессированных генов стержневого корня сахарной свеклы. Дж. Эксп. Бот. 53, 1533–1534. 10.1093/jexbot/53.373.1533 [PubMed] [CrossRef] [Google Scholar]
- Крепс Дж. А., Ву Ю. Дж., Чанг Х. С., Чжу Т., Ван С., Харпер Дж. Ф. (2002). Изменения транскриптома арабидопсиса в ответ на солевой, осмотический и холодовой стресс. Завод Физиол.
130, 2129–2141. 10.1104/pp.008532 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Куяла Т. С., Лопонен Дж. М., Клика К. Д., Пихлая К. (2000). Фенолы и бетацианины в корне красной свеклы ( Beta vulgaris ): распределение и влияние хранения в холодильнике на содержание общих фенолов и трех отдельных соединений. Дж. Агрик. Пищевая хим. 48, 5338–5342. 10.1021/jf000523q [PubMed] [CrossRef] [Google Scholar]
- Lake V., Olsson U., Willows R.D., Ханссон М. (2004). АТФазная активность субъединицы I хелатазы магния необходима для поддержания субъединицы D in vivo . Евро. Дж. Биохим. 271, 2182–2188. 10.1111/j.1432-1033.2004.04143.x [PubMed] [CrossRef] [Google Scholar]
- Ли Б., Хендерсон Д. А., Чжу Дж. (2005). Холодочувствительный транскриптом арабидопсиса и его регуляция с помощью ICE1. Растительная клетка 17, 3155–3175. 10.1105/tpc.105.035568 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Лер А., Кирш М. , Вирек Р., Шиманн Дж., Рауш Т. (1999). кДНК и геномное клонирование изоформ А и с субъединицы Н+-АТФазы V-типа сахарной свеклы: свидетельство координированной экспрессии во время развития растений и координированной индукции в ответ на сильное засоление. Завод Мол. биол.
39, 463–475. 10.1023/A:1006158310891 [PubMed] [CrossRef] [Google Scholar]
- Lein J.C., Asbach K., Tian Y., Schulte D., Li C., Koch G., et al. . (2007). Аналоги генов устойчивости сгруппированы на хромосоме 3 сахарной свеклы и косегрегируют с QTL устойчивости к ризомании.Геном 50, 61–71. 10.1139/G06-131 [PubMed] [CrossRef] [Google Scholar]
- Li H., Cao H., Wang Y., Pang Q., Ma C., Chen S. (2009). Протеомный анализ апомиктической моносомной линии присоединения сахарной свеклы М14. Дж. Протеомика 73, 297–308. 10.1016/j.jprot.2009.09.012 [PubMed] [CrossRef] [Google Scholar]
- Li H., Pan Y., Zhang Y., Wu C., Ma C., Yu B., et al. . (2015). Реакция мембранного протеома моносомной линии присоединения сахарной свеклы М14 на солевой стресс. Дж. Протеомика
127 (Пт А), 18–33. 10.1016/j.jprot.2015.03.025 [PubMed] [CrossRef] [Google Scholar]
- Li JL, Cui J., Cheng D.Y. (2015). Компьютерная идентификация и характеристика консервативных микроРНК и их генов-мишеней в свекле ( Beta vulgaris ). Жене. Мол. Рез. 14, 9103–9108. 10.4238/2015 [PubMed] [CrossRef] [Google Scholar]
- Li Q., Zhao P., Li J., Zhang C., Wang L., Ren Z. (2014). Полногеномный анализ семейства белков WD-повтора у огурца и арабидопсиса. Мол. Жене. Геномика 289, 103–124.10.1007/s00438-013-0789-x [PubMed] [CrossRef] [Google Scholar]
- Ликаузи Ф., Оме-Такаги М., Перата П. (2013). Фактор транскрипции APETALA2/Ethylene Responsive Factor (AP2/ERF): медиаторы реакции на стресс и программы развития. Новый Фитол. 199, 639–649. 10.1111/nph.12291 [PubMed] [CrossRef] [Google Scholar]
- Liu H., Stone SL (2013). Цитоплазматическая деградация фактора транскрипции Arabidopsis, нечувствительного к абсцизовой кислоте 5, опосредована лигазой E3 RING-типа KEEP ON GOING. Дж. Биол. хим.
288, 20267–20279. 10.1074/jbc.M113.465369 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Liu H., Wang Q., Yu M., Zhang Y., Wu Y., Zhang H. (2008) . Трансгенная солеустойчивая сахарная свекла ( Beta vulgaris L), конститутивно экспрессирующая вакуолярный Na/H-антипортер Arabidopsis thaliana , AtNHX3, накапливает больше растворимого сахара, но меньше соли в запасающих корнях. Окружающая среда растительной клетки. 31, 1325–1334. 10.1111/j.1365-3040.2008.01838.x [PubMed] [CrossRef] [Google Scholar]
- Liu L., Li Y., Li S., Hu N., He Y., Pong R., et al. . (2012). Сравнение систем секвенирования следующего поколения. Дж. Биомед. Биотехнолог. 2012:251364. 10.1155/2012/251364 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Liu Z., Li Y., Cao H., Ren D. (2015). Сравнительный фосфопротеомный анализ солечувствительных фосфопротеинов, регулируемых каскадом MKK9-MPK6 у арабидопсиса . Растениевод.
241, 138–150. 10.1016/j.plantsci.2015.10.005 [PubMed] [CrossRef] [Google Scholar]
- Llave C., Xie Z., Kasschau KD, Carrington JC (2002). Расщепление мишеней мРНК, похожей на чучело, направляется классом микроРНК арабидопсиса. Наука 297, 2053–2056 гг. 10.1126/science.1076311 [PubMed] [CrossRef] [Google Scholar]
- Luan Y., Cui J., Zhai J., Li J., Han L., Meng J. (2015). Высокопроизводительное секвенирование выявляет дифференциальную экспрессию миРНК в томате, инокулированном Phytophthora infestans. Планта 241, 1405–1416. 10.1007/s00425-015-2267-7 [PubMed] [CrossRef] [Google Scholar]
- Лукьянов С.А., Ребриков Д., Буздин А. А. (2007). Подавление субтрактивной гибридизации, в современных приложениях гибридизации нуклеиновых кислот, под ред. Буздин А. А., Лукьянов С. А. (Берлин: Springer; ), 53–84. [Google Scholar]
- Ma C., Wang Y., Wang Y., Wang L., Chen S., Li H. (2011). Идентификация гена бокса BvM14-MADS сахарной свеклы с помощью анализа дифференциальной экспрессии генов моносомной аддитивной линии M14. J. Физиол растений.
168, 1980–1986 гг. 10.1016/j.jplph.2011.05.027 [PubMed] [CrossRef] [Google Scholar]
- Мардис Э.Р. (2008). Влияние технологии секвенирования нового поколения на генетику. Тенденции Жене. 24, 133–141. 10.1016/j.tig.2007.12.007 [PubMed] [CrossRef] [Google Scholar]
- Миколайчик-Батор К., Блащик А., Чизнеевский М., Качицкий П. (2016). Характеристика и идентификация тритерпеновых сапонинов в корнях красной свеклы ( Beta vulgaris L) с использованием двух систем ВЭЖХ-МС. Пищевая хим. 192, 979–990. 10.1016/j.foodchem.2015.07.111 [PubMed] [CrossRef] [Google Scholar]
- Мишра К.Б., Яннаконе Р., Петроцца А., Мишра А., Арментано Н., Ла Веккья Г. и др. . (2012). Искусственная засухоустойчивость растений томатов отражается в эмиссии флуоресценции хлорофилла. Растениевод. 182, 79–86. 10.1016/j.plantsci.2011.03.022 [PubMed] [CrossRef] [Google Scholar]
- Митчелл Д. С., Ларами С. Э., Тереза Дж. Д. Р., Фредерик П. Б., Блэр Д. С., Тиффани М. Х. М. (2014). Транскрипционный ответ сои на обработку семян тиаметоксамом при наличии и отсутствии стресса, вызванного засухой.Геномика BMC
15:1055. 10.1186/1471-2164-15-1055 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Молитерни В.М., Пэрис Р., Онофри К., Орру Л., Каттивелли Л., Пасифико Д. и др. др. . (2015). Ранние изменения транскрипции у Beta vulgaris в ответ на низкую температуру. Планта 242, 187–201. 10.1007/s00425-015-2299-z [PubMed] [CrossRef] [Google Scholar]
- Moody L. A., Saidi Y., Smiles E. J., Bradshaw S. J., Meddings M., Winn P. J., et al. . (2012). Гомологи гена ARABIDILLO у растений базальной земли: видоспецифичная дупликация генов и вероятная функциональная избыточность.Планта 236, 1927–1941. 10.1007/s00425-012-1742-7 [PubMed] [CrossRef] [Google Scholar]
- Морено Д. А., Гарсия-Вигера К., Хил Дж., Хил-Искьердо А. (2008). Беталайны в эпоху глобальной агропродовольственной науки, технологий и здорового питания. Фитохим. преп.
7, 261–280. 10.1007/s11101-007-9084-y [CrossRef] [Google Scholar]
- Маннс Р., Тестер М. (2008). Механизмы солеустойчивости. Анну. Преподобный завод биол. 59, 651–681. 10.1146/annurev.arplant.59.032607.092911 [PubMed] [CrossRef] [Google Scholar]
- Ноглер Г.А. (1984). Гаметофитный апомиксис, в «Эмбриологии покрытосеменных растений», изд. Джохри Б.М. (Берлин: Springer;), 475–518. [Google Scholar]
- Оксман-Калдентей К. М., Сайто К. (2005). Интеграция геномики и метаболомики для разработки метаболических путей растений. Курс. мнение Биотехнолог. 16, 174–179. 10.1016/j.copbio.2005.02.007 [PubMed] [CrossRef] [Google Scholar]
- Озиас-Акинс П. (2006). Апомиксис: особенности развития и генетика. крит. Преподобный завод наук. 25, 199–214. 10.1080/07352680600563926 [CrossRef] [Google Scholar]
- Pinhero R., Пажекатту Р., Марангони А. Г., Лю К., Яда Р. Ю. (2011). Облегчение подслащивания картофеля при низкой температуре за счет экспрессии гена пируватдекарбоксилазы Arabidopsis и стресс-индуцируемого rd29A: предварительное исследование. Физиол. Мол. биол. Растения
17, 105–114. 10.1007/s12298-011-0056-8 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Паммер С., Данцлер В. Х., Лиен Ю. Х., Мокель Г. В., Фелькер К., Зильбернагл С. (2000). Реабсорбция бетаина в петлях Генле почек крыс in vivo .Являюсь. Дж. Физиол. почечная. Физиол. 278, 434–439. [PubMed] [Google Scholar]
- Puthoff DP, Smigocki AC (2007). Дифференциальная экспрессия корневых генов Beta vulgaris , вызванная кормлением насекомых, и их регуляция сигналами, связанными с защитой. Представитель растительных клеток. 26, 71–84. 10.1007/s00299-006-0201-yAN [PubMed] [CrossRef] [Google Scholar]
- Saad A.S.I., Li X., Li H.P., Huang T., Gao C.S., Guo M.W., et al. (2013). Ген NAC, чувствительный к стрессу риса, повышает устойчивость трансгенной пшеницы к засухе и солевому стрессу.Растениевод. 203–204, 33–40. 10.1016/j.plantsci.2012.12.016 [PubMed] [CrossRef] [Google Scholar]
- Сахеби М., Ханафи М. М. , Азизи П., Хаким А., Ашкани С., Абири Р. (2015). Подавление субтрактивной гибридизации по сравнению с секвенированием нового поколения в генной инженерии растений: проблемы и перспективы. Мол. Биотехнолог.
57, 880–903. 10.1007/s12033-015-9884-z [PubMed] [CrossRef] [Google Scholar]
- Сангер Ф., Никлен С., Коулсон А. Р. (1977). Секвенирование ДНК с помощью ингибиторов обрыва цепи.проц. Натл. акад. науч. США. 74, 5463–5467. 10.1073/pnas.74.12.5463 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Schuster S.C. (2008). Секвенирование нового поколения меняет современную биологию. Нац. Методы 5, 16–18. 10.1038/nmeth2156 [PubMed] [CrossRef] [Google Scholar]
- Setiawan A., Koch G., Barnes S.R., Jung C. (2000). Картирование локусов количественных признаков (QTL) для устойчивости к болезни пятнистости листьев Cercospora ( Cercospora beticola Sacc) у сахарной свеклы ( Beta vulgaris L.). Теор. заявл. Жене. 100, 1176–1182. 10.1007/s001220051421 [CrossRef] [Google Scholar]
- Shan Q. , Wang Y., Li J., Zhang Y., Chen K., Liang Z., et al. . (2013). Направленная модификация генома сельскохозяйственных растений с использованием системы CRISPR-Cas. Нац. Биотехнолог.
31, 686–688. 10.1038/nbt.2650 [PubMed] [CrossRef] [Google Scholar]
- Shinozaki K., Yamaguchi S.K. (2007). Генные сети, участвующие в реакции на засуху и устойчивости к стрессу. Дж. Эксп. Бот. 58, 221–227. 10.1093/jxb/erl164 [PubMed] [CrossRef] [Google Scholar]
- Сильва-Санчес С., Ли Х., Чен С. (2015). Последние достижения и проблемы в фосфопротеомике растений. протеомика 15, 1127–1141. 10.1002/pmic.201400410 [PubMed] [CrossRef] [Google Scholar]
- Stadermann K.B., Weisshaar B., Holtgräwe D. (2015). SMRT секвенирует только de novo сборки генома хлоропластов сахарной свеклы ( Beta vulgaris ). БМК Биоинформатика 16:295. 10.1186/s12859-015-0726-6 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Stintzing F.C., Carle R.(2007). Беталайны — новые перспективы для ученых-пищевиков. Тенденции Food Sci. Технол.
18, 514–525. 10.1016/j.tifs.2007.04.012 [CrossRef] [Google Scholar]
- Sun S., Yu J.P., Chen F., Zhao T.J., Fang X.H., Li Y.Q., et al. . (2008). TINY, чувствительный к дегидратации элемент (DRE), связывающий белок, подобный фактору транскрипции, соединяющий сигнальные пути, опосредованные DRE- и этилен-чувствительным элементом, у Arabidopsis. Дж. Биол. хим. 283, 6261–6271. 10.1074/jbc.M706800200 [PubMed] [CrossRef] [Google Scholar]
- Sunkar R., Чиннусами В., Чжу Дж., Чжу Дж. К. (2007). Малые РНК как крупные игроки в реакциях растений на абиотический стресс и лишение питательных веществ. Тенденции Растениевод. 12, 301–309. 10.1016/j.tplants.2007.05.001 [PubMed] [CrossRef] [Google Scholar]
- Тагучи К., Кубо Т., Такахаши Х., Абэ Х. (2011). Идентификация и точное картирование устойчивых QTL устойчивости к пятнистости листьев Cercospora у сахарной свеклы ( Beta vulgaris L). G3 (Бетесда) 1, 283–291. 10.1534/g3.111.000513 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Тагучи К. , Окадзаки К., Такахаши Х., Кубо Т., Миками Т. (2010). Молекулярное картирование гена, придающего устойчивость к корневой гнили Aphanomyces (черный корень) у сахарной свеклы ( Beta vulgaris L). Эуфитика
173, 409–418. 10.1007/s10681-010-0153-8 [CrossRef] [Google Scholar]
- Такабэ Т., Рай В., Хибино Т. (2006). Метаболическая инженерия глицинебетаина, Устойчивость растений к абиотическим стрессам, ред. Рай А.К., Такабе Т. (Берлин: Springer;), 137–151. [Google Scholar]
- Торрес М. А., Дангл Дж.Л. (2005). Функции оксидазы респираторного взрыва в биотических взаимодействиях, абиотическом стрессе и развитии. Курс. мнение биол. растений 8, 397–403. 10.1016/j.pbi.2005.05.014 [PubMed] [CrossRef] [Google Scholar]
- Влахонасиос К. Э., Томасов М. Ф., Тризенберг С. Дж. (2003). Нарушающие мутации транскрипционных генов-адаптеров ADA2b и GCN5 резко влияют на рост, развитие и экспрессию генов арабидопсиса. Растительная клетка 15, 626–638. 10.1105/tpc.007922 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Wakeel A. , Асиф А.Р., Питанн Б., Шуберт С. (2011). Анализ протеома сахарной свеклы ( Beta vulgaris L) выявил конститутивную адаптацию во время первой фазы солевого стресса. J. Физиол растений.
168, 519–526. 10.1016/j.jplph.2010.08.016 [PubMed] [CrossRef] [Google Scholar]
- Ван Ю. Н., Тан Л., Хоу Ю., Ван П., Ян Х., Вэй К. Л. (2016). Дифференциальный транскриптомный анализ листьев чайного растения ( Camellia sinensis ) дает всестороннее представление о защитных реакциях на атаку Ectropis oblique с использованием RNA-Seq.Функц. интегр. Геномика. [Epub перед печатью]. 10.1007/s10142-016-0491-2 [PubMed] [CrossRef] [Google Scholar]
- Wang Y., Zhan Y., Wu C., Gong S., Zhu N., Chen S., et al. (2012). Клонирование гена цистатина из сахарной свеклы М14, которое может повысить солеустойчивость растений. Растениевод. 191–192, 93–99. 10.1016/j.plantsci.2012.05.001 [PubMed] [CrossRef] [Google Scholar]
- Wu C., Ma C., Pan Y., Gong S., Zhao C., Chen S., et al. (2013). Ген глиоксалазы I сахарной свеклы М14 может повышать устойчивость растений к абиотическим стрессам.Дж. Плант Рез.
26, 415–425. 10.1007/s10265-012-0532-4 [PubMed] [CrossRef] [Google Scholar]
- Wybraniec S. (2005). Образование декарбоксилированных бетацианинов в нагретом корневом соке красной свеклы ( Betavulgaris L) проанализировано с помощью ЖХ-МС/МС. Дж. Агрик. Пищевая хим. 53, 3483–3487. 10.1021/jf048088d [PubMed] [CrossRef] [Google Scholar]
- Выбранец С., Сталица П., Спорна А., Немзер Б., Петшковски З., Михаловски Т. (2011). Антиоксидантная активность бетанидина: электрохимическое исследование в водной среде.Дж. Агрик. Пищевая хим. 59, 12163–12170. 10.1021/jf2024769 [PubMed] [CrossRef] [Google Scholar]
- Выбранец С., Старзак К., Скопиньска А., Немзер Б., Петшковский З., Михаловски Т. (2013). Изучение механизмов неферментативного окисления необетанина, бетанина и декарбоксилированных бетанинов. Дж. Агрик. Пищевая хим. 61, 6465–6476. 10.1021/jf400818s [PubMed] [CrossRef] [Google Scholar]
- Xu J. , Lan H., Fang H., Huang X., Zhang H., Huang J. (2015). Количественный протеомный анализ реакции риса ( Oryza sativa L) на соль.ПЛОС ОДИН
10:e0120978. 10.1371/journal.pone.0120978 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Ямада Н., Промден В., Ямане К., Тамагаке Х., Хибино Т., Танака Ю. и др. . . (2009). Преимущественное накопление бетаина, не связанного с холинмонооксигеназой, в молодых листьях сахарной свеклы – важность транслокации бетаина на большие расстояния в нормальных условиях и в условиях солевого стресса. J. Физиол растений. 166, 2058–2070 гг. 10.1016/j.jplph.2009.06.016 [PubMed] [CrossRef] [Google Scholar]
- Ян Л., Ма С., Ван Л., Чен С., Ли Х. (2012). Вызванные солевым стрессом изменения протеома и транскриптома в моносомной линии добавления сахарной свеклы М14. J. Физиол растений. 169, 839–850. 10.1016/j.jplph.2012.01.023 [PubMed] [CrossRef] [Google Scholar]
- Yang L., Zhang Y., Zhu N., Koh J., Ma C., Pan Y., et al. . (2013). Протеомный анализ солеустойчивости моносомной линии присоединения сахарной свеклы М14. Дж. Протеом Рез.
12, 4931–4950. 10.1021/pr400177m [PubMed] [CrossRef] [Google Scholar]
- Инь Ф., Гао Дж., Лю М., Цинь С., Чжан В., Ян А. и др. . (2014). Полногеномный анализ профиля экспрессии микроРНК, реагирующей на водный стресс, в корнях табака. Функц. интегр. Геномика 14, 319–332. 10.1007/s10142-014-0365-4 [PubMed] [CrossRef] [Google Scholar]
- Йолджу С., Оздемир Ф., Гюлер А., Бор М. (2016). Ацетилирование гистонов влияет на активацию транскрипции POX у Beta vulgaris L. и Beta maritima L. при солевом стрессе. Завод Физиол. Биохим. 100, 37–46. 10.1016/Дж.plaphy.2015.12.019 [PubMed] [CrossRef] [Google Scholar]
- Zhang Y., Lai J., Sun S., Li Y., Liu Y., Liang L., et al. . (2008). Сравнительный анализ транскриптов галофита Thellungiella halophila . Дж. Интегр. биол. растений 50, 1327–1335. 10.1111/j.1744-7909.2008.00740.x [PubMed] [CrossRef] [Google Scholar]
- Zhu H. , Bi YD, Yu LJ, Guo D.D., Wang B.C. (2009). Сравнительный протеомный анализ апомиктической моносомной линии присоединения Beta corolliflora и Beta vulgaris L.в сахарной свекле. Мол. биол. Респ.
36, 2093–2098. 10.1007/s11033-008-9421-2 [PubMed] [CrossRef] [Google Scholar]
 Окружающая среда растительной клетки.
31, 11–38. 10.1111/j.1365-3040.2007.01727.x [PubMed] [CrossRef] [Google Scholar]
Окружающая среда растительной клетки.
31, 11–38. 10.1111/j.1365-3040.2007.01727.x [PubMed] [CrossRef] [Google Scholar] Х., Хамада А., Ямада Н., Рай В., Хибино Т., Такабэ Т. (2007). Регуляция синтеза бетаина с помощью предшественника и экспрессии холинмонооксигеназы в Amaranthus tricolor.Дж. Эксп. Бот.
58, 4203–4212. 10.1093/jxb/erm278 [PubMed] [CrossRef] [Google Scholar]
Х., Хамада А., Ямада Н., Рай В., Хибино Т., Такабэ Т. (2007). Регуляция синтеза бетаина с помощью предшественника и экспрессии холинмонооксигеназы в Amaranthus tricolor.Дж. Эксп. Бот.
58, 4203–4212. 10.1093/jxb/erm278 [PubMed] [CrossRef] [Google Scholar] 1007/s10142-015-0451-2 [PubMed] [CrossRef] [Google Scholar]
1007/s10142-015-0451-2 [PubMed] [CrossRef] [Google Scholar] Растительная клетка
25, 1158–1173. 10.1105/tpc.113.109942 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Растительная клетка
25, 1158–1173. 10.1105/tpc.113.109942 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Р., Эльбешбиши Э., Хафез Х., Ли Х. С. (2015). Производство водорода из сока сахарной свеклы с использованием интегрированного биоводородного процесса темного брожения и микробного электролизера. Биоресурсная технология.
198, 223–230. 10.1016/j.biortech.2015.08.048 [PubMed] [CrossRef] [Google Scholar]
Р., Эльбешбиши Э., Хафез Х., Ли Х. С. (2015). Производство водорода из сока сахарной свеклы с использованием интегрированного биоводородного процесса темного брожения и микробного электролизера. Биоресурсная технология.
198, 223–230. 10.1016/j.biortech.2015.08.048 [PubMed] [CrossRef] [Google Scholar] З., Будак Х. (2009). Секвенирование более 13 000 экспрессированных меток последовательностей из шести субтрактивных библиотек кДНК диких и современных пшениц после медленного засушливого стресса. Окружающая среда растительной клетки.
32, 220–236. 10.1111/j.1365-3040.2008.01915.x [PubMed] [CrossRef] [Google Scholar]
З., Будак Х. (2009). Секвенирование более 13 000 экспрессированных меток последовательностей из шести субтрактивных библиотек кДНК диких и современных пшениц после медленного засушливого стресса. Окружающая среда растительной клетки.
32, 220–236. 10.1111/j.1365-3040.2008.01915.x [PubMed] [CrossRef] [Google Scholar]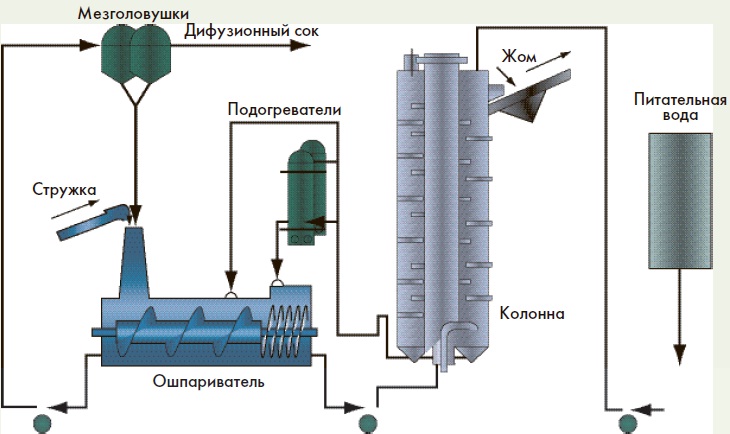 R., Trethewey R. N., Krotzky A. J., Willmitzer L.(2004). Метаболитное профилирование: от диагностики к системной биологии. Нац. Преподобный Мол. Клеточная биол.
5, 763–769. 10.1038/nrm1451 [PubMed] [CrossRef] [Google Scholar]
R., Trethewey R. N., Krotzky A. J., Willmitzer L.(2004). Метаболитное профилирование: от диагностики к системной биологии. Нац. Преподобный Мол. Клеточная биол.
5, 763–769. 10.1038/nrm1451 [PubMed] [CrossRef] [Google Scholar] Карта привязанного сцепления для сахарной свеклы на основе маркеров AFLP, SNP и RAPD и картирование QTL нового источника устойчивости к вирусу некроза желтой жилки свеклы.Теор. заявл. Жене.
114, 1151–1160. 10.1007/s00122-007-0507-3 [PubMed] [CrossRef] [Google Scholar]
Карта привязанного сцепления для сахарной свеклы на основе маркеров AFLP, SNP и RAPD и картирование QTL нового источника устойчивости к вирусу некроза желтой жилки свеклы.Теор. заявл. Жене.
114, 1151–1160. 10.1007/s00122-007-0507-3 [PubMed] [CrossRef] [Google Scholar] 200401101 [PubMed] [CrossRef] [Google Scholar]
200401101 [PubMed] [CrossRef] [Google Scholar] , Россо Ф., Штайнрюкен Г., Беккерса Р. (Бельгия: Международный институт исследований свеклы; ), 5–16. [Google Scholar]
, Россо Ф., Штайнрюкен Г., Беккерса Р. (Бельгия: Международный институт исследований свеклы; ), 5–16. [Google Scholar]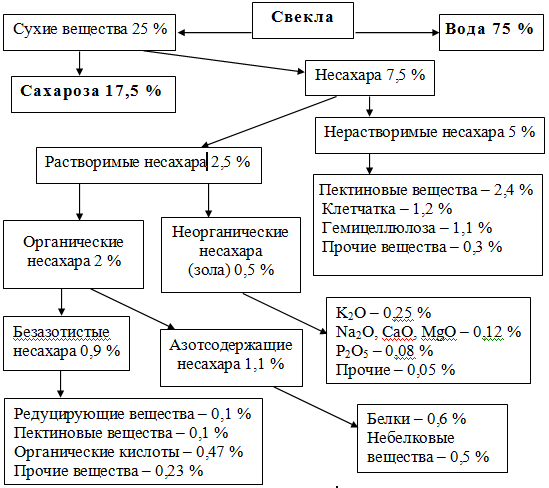 T., Thomas-Oates J. и соавт. . (2015). Метаболомика растений на основе масс-спектрометрии: реакция метаболитов на абиотический стресс. Масс-спектр. Ред. [Epub перед печатью]. 10.1002/mas.21449 [PubMed] [CrossRef] [Google Scholar]
T., Thomas-Oates J. и соавт. . (2015). Метаболомика растений на основе масс-спектрометрии: реакция метаболитов на абиотический стресс. Масс-спектр. Ред. [Epub перед печатью]. 10.1002/mas.21449 [PubMed] [CrossRef] [Google Scholar] Завод Физиол.
130, 2129–2141. 10.1104/pp.008532 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Завод Физиол.
130, 2129–2141. 10.1104/pp.008532 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] , Вирек Р., Шиманн Дж., Рауш Т. (1999). кДНК и геномное клонирование изоформ А и с субъединицы Н+-АТФазы V-типа сахарной свеклы: свидетельство координированной экспрессии во время развития растений и координированной индукции в ответ на сильное засоление. Завод Мол. биол.
39, 463–475. 10.1023/A:1006158310891 [PubMed] [CrossRef] [Google Scholar]
, Вирек Р., Шиманн Дж., Рауш Т. (1999). кДНК и геномное клонирование изоформ А и с субъединицы Н+-АТФазы V-типа сахарной свеклы: свидетельство координированной экспрессии во время развития растений и координированной индукции в ответ на сильное засоление. Завод Мол. биол.
39, 463–475. 10.1023/A:1006158310891 [PubMed] [CrossRef] [Google Scholar] Дж. Протеомика
127 (Пт А), 18–33. 10.1016/j.jprot.2015.03.025 [PubMed] [CrossRef] [Google Scholar]
Дж. Протеомика
127 (Пт А), 18–33. 10.1016/j.jprot.2015.03.025 [PubMed] [CrossRef] [Google Scholar] Дж. Биол. хим.
288, 20267–20279. 10.1074/jbc.M113.465369 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж. Биол. хим.
288, 20267–20279. 10.1074/jbc.M113.465369 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 10.1016/j.plantsci.2015.10.005 [PubMed] [CrossRef] [Google Scholar]
10.1016/j.plantsci.2015.10.005 [PubMed] [CrossRef] [Google Scholar] J. Физиол растений.
168, 1980–1986 гг. 10.1016/j.jplph.2011.05.027 [PubMed] [CrossRef] [Google Scholar]
J. Физиол растений.
168, 1980–1986 гг. 10.1016/j.jplph.2011.05.027 [PubMed] [CrossRef] [Google Scholar] Х. М. (2014). Транскрипционный ответ сои на обработку семян тиаметоксамом при наличии и отсутствии стресса, вызванного засухой.Геномика BMC
15:1055. 10.1186/1471-2164-15-1055 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Х. М. (2014). Транскрипционный ответ сои на обработку семян тиаметоксамом при наличии и отсутствии стресса, вызванного засухой.Геномика BMC
15:1055. 10.1186/1471-2164-15-1055 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] преп.
7, 261–280. 10.1007/s11101-007-9084-y [CrossRef] [Google Scholar]
преп.
7, 261–280. 10.1007/s11101-007-9084-y [CrossRef] [Google Scholar] Физиол. Мол. биол. Растения
17, 105–114. 10.1007/s12298-011-0056-8 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Физиол. Мол. биол. Растения
17, 105–114. 10.1007/s12298-011-0056-8 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] , Азизи П., Хаким А., Ашкани С., Абири Р. (2015). Подавление субтрактивной гибридизации по сравнению с секвенированием нового поколения в генной инженерии растений: проблемы и перспективы. Мол. Биотехнолог.
57, 880–903. 10.1007/s12033-015-9884-z [PubMed] [CrossRef] [Google Scholar]
, Азизи П., Хаким А., Ашкани С., Абири Р. (2015). Подавление субтрактивной гибридизации по сравнению с секвенированием нового поколения в генной инженерии растений: проблемы и перспективы. Мол. Биотехнолог.
57, 880–903. 10.1007/s12033-015-9884-z [PubMed] [CrossRef] [Google Scholar] , Wang Y., Li J., Zhang Y., Chen K., Liang Z., et al. . (2013). Направленная модификация генома сельскохозяйственных растений с использованием системы CRISPR-Cas. Нац. Биотехнолог.
31, 686–688. 10.1038/nbt.2650 [PubMed] [CrossRef] [Google Scholar]
, Wang Y., Li J., Zhang Y., Chen K., Liang Z., et al. . (2013). Направленная модификация генома сельскохозяйственных растений с использованием системы CRISPR-Cas. Нац. Биотехнолог.
31, 686–688. 10.1038/nbt.2650 [PubMed] [CrossRef] [Google Scholar] Тенденции Food Sci. Технол.
18, 514–525. 10.1016/j.tifs.2007.04.012 [CrossRef] [Google Scholar]
Тенденции Food Sci. Технол.
18, 514–525. 10.1016/j.tifs.2007.04.012 [CrossRef] [Google Scholar] , Окадзаки К., Такахаши Х., Кубо Т., Миками Т. (2010). Молекулярное картирование гена, придающего устойчивость к корневой гнили Aphanomyces (черный корень) у сахарной свеклы ( Beta vulgaris L). Эуфитика
173, 409–418. 10.1007/s10681-010-0153-8 [CrossRef] [Google Scholar]
, Окадзаки К., Такахаши Х., Кубо Т., Миками Т. (2010). Молекулярное картирование гена, придающего устойчивость к корневой гнили Aphanomyces (черный корень) у сахарной свеклы ( Beta vulgaris L). Эуфитика
173, 409–418. 10.1007/s10681-010-0153-8 [CrossRef] [Google Scholar] , Асиф А.Р., Питанн Б., Шуберт С. (2011). Анализ протеома сахарной свеклы ( Beta vulgaris L) выявил конститутивную адаптацию во время первой фазы солевого стресса. J. Физиол растений.
168, 519–526. 10.1016/j.jplph.2010.08.016 [PubMed] [CrossRef] [Google Scholar]
, Асиф А.Р., Питанн Б., Шуберт С. (2011). Анализ протеома сахарной свеклы ( Beta vulgaris L) выявил конститутивную адаптацию во время первой фазы солевого стресса. J. Физиол растений.
168, 519–526. 10.1016/j.jplph.2010.08.016 [PubMed] [CrossRef] [Google Scholar] (2013). Ген глиоксалазы I сахарной свеклы М14 может повышать устойчивость растений к абиотическим стрессам.Дж. Плант Рез.
26, 415–425. 10.1007/s10265-012-0532-4 [PubMed] [CrossRef] [Google Scholar]
(2013). Ген глиоксалазы I сахарной свеклы М14 может повышать устойчивость растений к абиотическим стрессам.Дж. Плант Рез.
26, 415–425. 10.1007/s10265-012-0532-4 [PubMed] [CrossRef] [Google Scholar] , Lan H., Fang H., Huang X., Zhang H., Huang J. (2015). Количественный протеомный анализ реакции риса ( Oryza sativa L) на соль.ПЛОС ОДИН
10:e0120978. 10.1371/journal.pone.0120978 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Lan H., Fang H., Huang X., Zhang H., Huang J. (2015). Количественный протеомный анализ реакции риса ( Oryza sativa L) на соль.ПЛОС ОДИН
10:e0120978. 10.1371/journal.pone.0120978 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] (2013). Протеомный анализ солеустойчивости моносомной линии присоединения сахарной свеклы М14. Дж. Протеом Рез.
12, 4931–4950. 10.1021/pr400177m [PubMed] [CrossRef] [Google Scholar]
(2013). Протеомный анализ солеустойчивости моносомной линии присоединения сахарной свеклы М14. Дж. Протеом Рез.
12, 4931–4950. 10.1021/pr400177m [PubMed] [CrossRef] [Google Scholar] , Bi YD, Yu LJ, Guo D.D., Wang B.C. (2009). Сравнительный протеомный анализ апомиктической моносомной линии присоединения Beta corolliflora и Beta vulgaris L.в сахарной свекле. Мол. биол. Респ.
36, 2093–2098. 10.1007/s11033-008-9421-2 [PubMed] [CrossRef] [Google Scholar]
, Bi YD, Yu LJ, Guo D.D., Wang B.C. (2009). Сравнительный протеомный анализ апомиктической моносомной линии присоединения Beta corolliflora и Beta vulgaris L.в сахарной свекле. Мол. биол. Респ.
36, 2093–2098. 10.1007/s11033-008-9421-2 [PubMed] [CrossRef] [Google Scholar]Сахарная свекла — обзор
C. Сахарная свекла (
Beta vulgaris ) Сахарная свекла выращивается для производства сахара с тех пор середины 18 века в Восточной Европе и в настоящее время является ценной традиционной культурой в регионах с умеренным климатом. Около 37% мирового сахара производится из свеклы (остальная часть добывается из сахарного тростника), и ежегодно из свеклы производится около 40 млн тонн сахара (Winner, 1993).В настоящее время сахарная свекла редко выращивается в органических системах из-за высоких затрат на прополку и отсутствия рынков сбыта и органически сертифицированных заводов по переработке (Lampkin, 1990), хотя в настоящее время рынок доступен в Великобритании (Lampkin and Measures, 2001) и некоторые другие страны.
Как и другие незерновые культуры, сахарная свекла может обеспечить ценную передышку от целого ряда болезней и вредителей зерновых. Однако его не следует выращивать в том же севообороте, что и масличный рапс, так как он страдает от общих проблем с вредителями и болезнями, включая свекольную нематоду и альтернариоз (Agrios, 1997; Cooke, 1993; Duffus and Ruppel, 1993).Также необходим значительный перерыв между последующими посевами сахарной свеклы, чтобы избежать зарастания сорной свеклой, семена которой могут сохраняться в почве в течение нескольких лет.
Урожай свеклы потенциально может оказать положительное влияние на борьбу с сорняками за счет необходимости и возможности борьбы с сорняками (Лампкин, 1990). Однако из-за относительно низкой ценности урожая борьба с сорняками проводится только для предотвращения потерь урожая из-за конкуренции, а популяции сорняков во многих случаях могут быть выше после посева сахарной свеклы, чем до него (Halder, pers.комм.).
Сахарная свекла, как правило, высевается сеялкой-шведкой, и требуется специальная техника для уборки корней, чтобы подрезать и подрезать свеклу во время или до уборки (Bray and Thompson, 1985).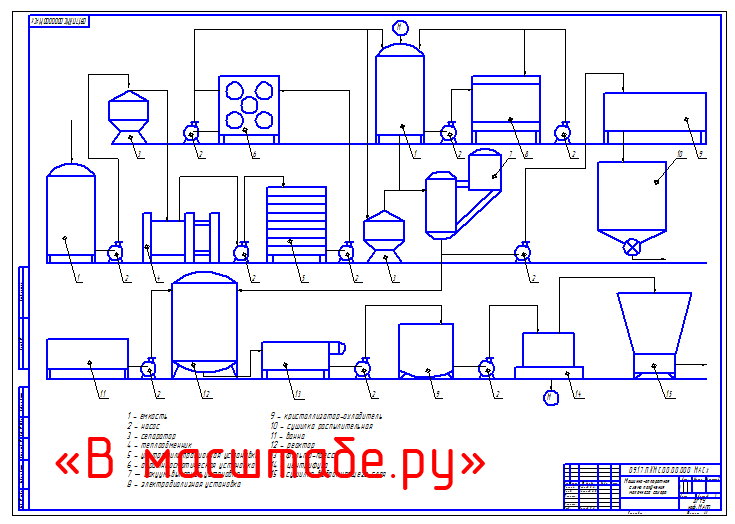 Культура имеет низкую потребность в азоте, а высокие остаточные концентрации азота в почве могут снизить содержание сахара в свекле. Поэтому его следует выращивать после зерновых или требовательных к азоту культур (Лампкин, 1990). Если сахарную свеклу выращивают перед озимыми зерновыми, нормальная дата уборки свеклы будет после идеальной даты посева зерновых.Может оказаться необходимым, особенно на тяжелых почвах, рано убрать свеклу. Это привело бы к снижению урожайности, но позволило бы сеять раньше и, следовательно, лучше укоренить более прибыльную культуру озимых зерновых (Bray and Thompson, 1985). Урожай свеклы, посеянный после злака, также может вызвать проблемы в севообороте. Зерновые обычно собирают в июле/августе, а свеклу сеют только в марте/апреле следующего года. Это оставляет почву голой примерно на 8 месяцев в зимний период, что делает нитраты восприимчивыми к выщелачиванию (Allison et al., 1996). Риск эрозии почвы также увеличивается на залежных почвах, особенно зимой.
Культура имеет низкую потребность в азоте, а высокие остаточные концентрации азота в почве могут снизить содержание сахара в свекле. Поэтому его следует выращивать после зерновых или требовательных к азоту культур (Лампкин, 1990). Если сахарную свеклу выращивают перед озимыми зерновыми, нормальная дата уборки свеклы будет после идеальной даты посева зерновых.Может оказаться необходимым, особенно на тяжелых почвах, рано убрать свеклу. Это привело бы к снижению урожайности, но позволило бы сеять раньше и, следовательно, лучше укоренить более прибыльную культуру озимых зерновых (Bray and Thompson, 1985). Урожай свеклы, посеянный после злака, также может вызвать проблемы в севообороте. Зерновые обычно собирают в июле/августе, а свеклу сеют только в марте/апреле следующего года. Это оставляет почву голой примерно на 8 месяцев в зимний период, что делает нитраты восприимчивыми к выщелачиванию (Allison et al., 1996). Риск эрозии почвы также увеличивается на залежных почвах, особенно зимой. Органические фермеры часто предпочитают выращивать покровную культуру, такую как пастбищная рожь или озимая вика, вместо зимы между культурами, которые собирают рано, и культурами, которые сажают следующей весной. Такие культуры снижают вероятность и серьезность эрозии почвы и выщелачивания нитратов, и их можно вспахивать перед посевом следующей культуры (Soil Association, 1998).
Органические фермеры часто предпочитают выращивать покровную культуру, такую как пастбищная рожь или озимая вика, вместо зимы между культурами, которые собирают рано, и культурами, которые сажают следующей весной. Такие культуры снижают вероятность и серьезность эрозии почвы и выщелачивания нитратов, и их можно вспахивать перед посевом следующей культуры (Soil Association, 1998).
Когда кроны и ботва посевов сахарной свеклы вспахивают обратно в почву после сбора урожая, добавляется значительное количество органических веществ и питательных веществ.Например, урожай сахарной свеклы может внести в почву 0,6–1,01 га -1 сухого вещества после заделки пожнивных остатков (Lampkin, 1990). Брей и Томпсон (1985) подсчитали, что 105 кг га 90 746 -1 90 747 N, 35 кг га 90 746 -1 90 747 P и 145 кг га 90 746 -1 90 747 K были возвращены в почву из крон и верхушек 50-тонного га −1 урожая сахарной свеклы. Пшенице, посеянной после сахарной свеклы, требовалось меньше азота при заделке ботвы, чем после пшеницы (Sylvester-Bradley and Shepherd, 1997). Однако азот, высвобождаемый из инкорпорированной крошки, подвержен потерям в результате выщелачивания, денитрификации или улетучивания (Sylvester-Bradley and Shepherd, 1997).
Однако азот, высвобождаемый из инкорпорированной крошки, подвержен потерям в результате выщелачивания, денитрификации или улетучивания (Sylvester-Bradley and Shepherd, 1997).
В Европе существует небольшой, но растущий рынок для этой культуры. До недавнего времени в Великобритании потребность в перерабатывающих мощностях ограничивала интерес к культуре, которую можно выращивать только по контракту. Без ценовых надбавок сахарная свекла имеет разумную валовую прибыль, сравнимую с полевыми бобами и озимым овсом (таблица X), но после испытаний в 2000 г. British Sugar plc предлагает контракты на 2001 г. с надбавкой к цене, в результате чего валовая прибыль достигает 1799 фунтов стерлингов на га −1 .Однако затраты на полевые операции снижают чистую прибыль ниже, чем у озимого овса. Спрос на органический сахар, особенно для переработки, растет, и мощности по переработке, вероятно, возрастут. В настоящее время в сельском хозяйстве Великобритании или Европы не существует субсидий на производство сахарной свеклы. В Европе существует небольшой, но растущий рынок для этой культуры.
В Европе существует небольшой, но растущий рынок для этой культуры.
Как перерабатывается сахар — IFT.org
Закрыть расширятьИзмельченная сахарная свекла движется по конвейерной ленте на пути к переработке.© Avatar_023/iStock/Getty Images Plus
Колонка этого месяца «Обработка» продолжает тему «Как это обрабатывается?» с упором на сахар. Помимо изучения истории, рынка и потребления сахара, в колонке будут описаны этапы обработки и производства.
История, рынок и потребление Сахар получают из двух основных растительных источников: сахарного тростника и сахарной свеклы. Как один из старейших товаров в мире, он был впервые одомашнен около 8000 г. до н.э.C. коренными жителями Новой Гвинеи. Из-за своей ценности сахар часто хранили под замком в сахарных сейфах. В 1751 году в Луизиане был посажен первый в США сахарный тростник, что положило начало сахарной промышленности США, которая сегодня обеспечивает 142 000 рабочих мест в 22 штатах.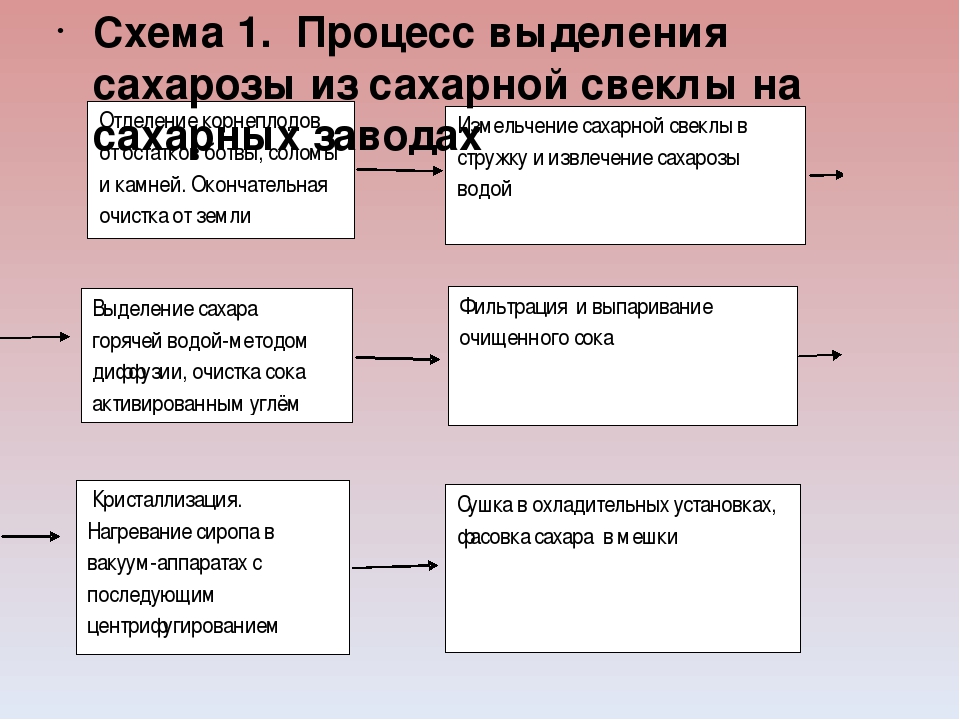
производится более чем в 120 странах мира; почти 80% сахара поступает из сахарного тростника, а остальная часть — из сахарной свеклы. В Соединенных Штатах 55 % производства сахара приходится на сахарную свеклу и 45 % — на сахарный тростник.
По данным Сельскохозяйственной службы Министерства сельского хозяйства США (USDA), мировое производство сахара в этом году достигнет 188 миллионов метрических тонн, что на 22 миллиона тонн больше, чем в прошлом году. Бразилия (39,5 млн тонн) в настоящее время является крупнейшим производителем сахара, за ней следуют Индия (33,7 млн тонн) и Европейский Союз (17,7 млн тонн). Прогнозируется, что в этом году Соединенные Штаты произведут 8,2 миллиона тонн сахара.
Сахарпроизводится более чем в 120 странах мира; почти 80% сахара поступает из сахарного тростника, а остальная часть — из сахарной свеклы.
За последние 20 лет потребление сахара в США снизилось. По данным Сахарной ассоциации, годовое потребление сахара составляло 89,6 фунтов на человека в 1999 году; в 2016 году он составлял 77,1 фунта на человека, что эквивалентно примерно 16 чайным ложкам в день.
По данным Сахарной ассоциации, годовое потребление сахара составляло 89,6 фунтов на человека в 1999 году; в 2016 году он составлял 77,1 фунта на человека, что эквивалентно примерно 16 чайным ложкам в день.
Сахар бывает самых разных форм. Наиболее распространенным является сахар-песок, также называемый столовым сахаром, который является наиболее рафинированной формой. Сахарный песок с более крупными кристаллами называется грубым сахаром; с более мелкими кристаллами его называют сверхтонким сахаром.Жемчужный сахар состоит из конгломератов сахарных частиц. Коричневый сахар в различных цветовых диапазонах определяется количеством присутствующей в них патоки; они варьируются от ярко-желтого до темно-коричневого. Специальные типы сахара включают сахарную пудру (содержит кукурузный крахмал), сахар типа демерара (светло-коричневый сахар с крупными кристаллами, покрытыми патокой), сахар мусковадо (более липкий, грубый коричневый сахар, получаемый на ранней стадии рафинирования) и сахар типа турбинадо ( частично очищенный, полурафинированный коричневый сахар, также называемый Сахар-сырец . Настоящий сахар-сырец относится к частично очищенному сахару, который является мягким и коричневым и содержит примеси.
Настоящий сахар-сырец относится к частично очищенному сахару, который является мягким и коричневым и содержит примеси.
• Сбор урожая. Сахарный тростник и сахарная свекла обычно убираются с полей механическим способом. Собранное растительное сырье часто просеивают для удаления грязи и камней перед погрузкой на грузовики и доставкой на завод для обработки.
Закрыть расширятьЖелезнодорожные бункеры, загруженные сырым сахарным тростником, ждут на складе, прежде чем отправиться на мельницу для дробления и переработки.© BeyondImages/iStock/Getty Images Plus
• Мойка и начальная подготовка. Как только сахарный тростник или сахарная свекла поступают на завод, их тщательно промывают. Промывка может происходить на лентах, которые обрызгиваются водой, или в дымоходах, полных воды. Вращающиеся барабаны часто используются в качестве моечных станций. Вода распыляется в барабан, и продукт вращается внутри барабана, трется о себя, удаляя грязь. После промывки сахарный тростник или сахарная свекла транспортируются на завод с помощью шнеков или ремней.Сахарный тростник измельчается с помощью измельчителей с поворотным молотком или дробильных вальцов с большими канавками. Сахарную свеклу нарезают с помощью машин для нарезки, которые рвут ее на полоски меньшего размера, чем картофель фри, называемые косетками. Косетки замачивают в баках с горячей водой, а измельченный сахарный тростник опрыскивают горячей водой. Оба процесса служат для набухания растительных клеток при подготовке к экстракции.
Вода распыляется в барабан, и продукт вращается внутри барабана, трется о себя, удаляя грязь. После промывки сахарный тростник или сахарная свекла транспортируются на завод с помощью шнеков или ремней.Сахарный тростник измельчается с помощью измельчителей с поворотным молотком или дробильных вальцов с большими канавками. Сахарную свеклу нарезают с помощью машин для нарезки, которые рвут ее на полоски меньшего размера, чем картофель фри, называемые косетками. Косетки замачивают в баках с горячей водой, а измельченный сахарный тростник опрыскивают горячей водой. Оба процесса служат для набухания растительных клеток при подготовке к экстракции.
• Извлечение сока. Сахарная стружка закачивается на дно резервуаров высотой от 10 до 20 метров.Вращающийся вал в центре перемещает полосы сахарной свеклы вверх против нисходящего потока воды по мере извлечения сахара. Из сахарного тростника измельчение используется для извлечения сока. Серия из пяти мельниц сжимает волокна сахарного тростника и отделяет сок от багассы, которую позже можно использовать в качестве источника топлива.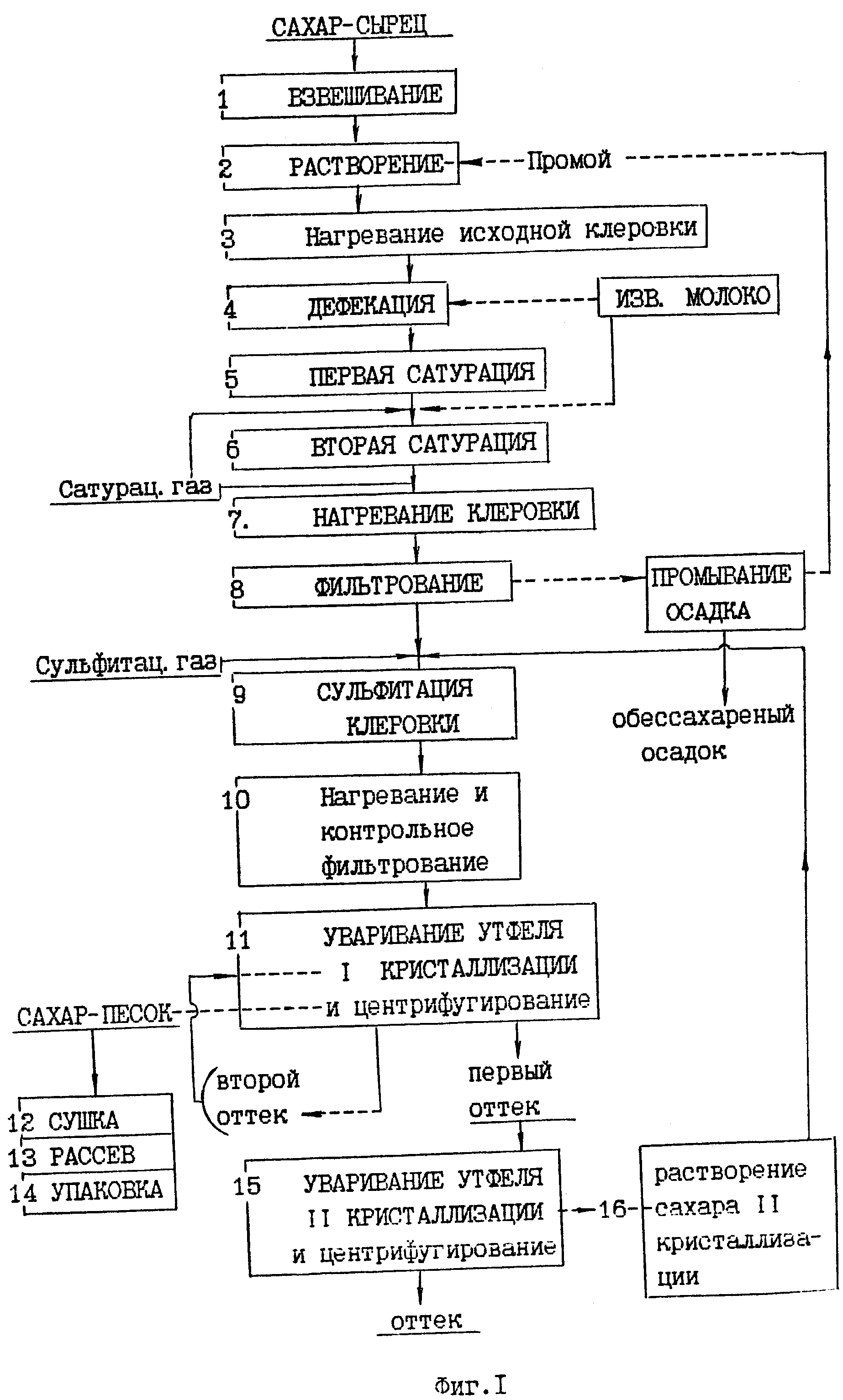 Исходный сок темно-зеленого цвета, кислый и мутный. Сок собирают в большие чаны и измеряют концентрацию сахара.
Исходный сок темно-зеленого цвета, кислый и мутный. Сок собирают в большие чаны и измеряют концентрацию сахара.
• Очистка сока. Высокие башни высотой от 10 до 20 метров используются для очистки и осветления тростникового сока. Сок вводят в верхнюю часть колонны, а пары диоксида серы вводят в нижнюю часть. Диоксид серы поднимается вверх через колонну в процессе, называемом сульфитацией. Газ добавляется в количестве от 120 до 200 фунтов на миллион фунтов сока. Карбонизация или подщелачивание используются для дальнейшего отделения растворимых несахарных материалов от сахарного сока. Карбонизация использует карбонат кальция или сульфит кальция, чтобы способствовать осаждению.Сок нагревают, чтобы денатурировать содержание белка, а затем смешивают с суспензией гидроксида кальция, называемой известковым молоком. На этом этапе также могут быть введены пузырьки углекислого газа для снижения щелочности и осаждения шлама сатурации, который фильтруется для очистки сока.
Процесс очистки обычно занимает несколько часов. В конце ил удаляется со дна резервуара, а сок удаляется сверху. Вторичная фильтрация используется для извлечения остаточного сахара из осадка.При этом образуется материал, называемый грязью, который позже можно использовать для удобрения полей.
Затем осветленный сок кипятят в нескольких вакуумных испарителях до тех пор, пока он не достигнет концентрации 50–65% сахара. Каждый последующий испаритель в этой серии имеет более высокое вакуумметрическое давление, чем предыдущий испаритель, что приводит к кипению сахарного сиропа при все более низких температурах по мере его прохождения в процессе. Осадок снимается с верхней части испарителей с помощью лопастных скиммеров, в результате чего получается густой, почти бесцветный сахарный сироп.
• Кристаллизация. На следующем этапе производственного процесса используется одноступенчатый вакуумный лоток для выпаривания сиропа до его насыщения кристаллами сахара, которые образуются в процессе, называемом засевом. Млечный раствор чистой сахарозы, суспендированной в спирте и глицерине, представляет собой затравку, которую медленно добавляют в сироп. Маленькие крупинки сахара, присутствующие в растворе, служат ядрами, помогая вытягивать сахар из раствора и превращать его в кристаллы.Когда смесь кипятится в вакуумной посуде, вода испаряется, а кристаллы сахара продолжают превращаться в пасту, называемую утфелем, густую смесь сиропа и кристаллов сахара. Затем эту смесь переносят в большую емкость, называемую кристаллизатором, где утфель медленно перемешивают и охлаждают, продолжая процесс кристаллизации.
Млечный раствор чистой сахарозы, суспендированной в спирте и глицерине, представляет собой затравку, которую медленно добавляют в сироп. Маленькие крупинки сахара, присутствующие в растворе, служат ядрами, помогая вытягивать сахар из раствора и превращать его в кристаллы.Когда смесь кипятится в вакуумной посуде, вода испаряется, а кристаллы сахара продолжают превращаться в пасту, называемую утфелем, густую смесь сиропа и кристаллов сахара. Затем эту смесь переносят в большую емкость, называемую кристаллизатором, где утфель медленно перемешивают и охлаждают, продолжая процесс кристаллизации.
• Центрифугирование. Для разделения утфеля на кристаллы сахара и патоку утфель добавляют в высокоскоростную центрифугу.Центрифуга, вращающаяся со скоростью от 1000 до 2800 оборотов в минуту, содержит перфорированную металлическую цилиндрическую корзину. Во время центрифугирования меласса выходит из футерованной корзины центрифуги и вытягивается наружу центрифуги, где ее удаляют и отправляют в резервуары для хранения.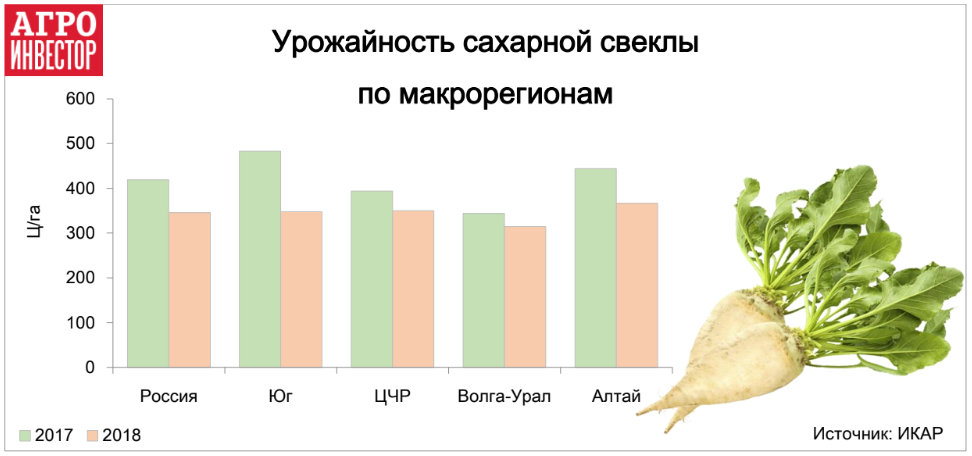 Сахар остается в футерованной корзине центрифуги. Родниковая вода используется для промывки кристаллов по мере их центрифугирования.
Сахар остается в футерованной корзине центрифуги. Родниковая вода используется для промывки кристаллов по мере их центрифугирования.
• Сушка и упаковка. Влажные кристаллы сахара сушат в больших сушилках с горячим воздухом, достигая содержания влаги до 0.02%. Затем сахар осторожно перемалывается через нагретый воздух в грануляторе. Затем высушенные кристаллы разделяются на разные размеры с помощью вибрационных сит и помещаются в бункеры для хранения. Наконец, сахар упаковывается для потребителя.
Будущее сахараСахар — универсальный и важный пищевой ингредиент с широким спектром преимуществ. Он не только придает продуктам сладость и вкус, но также добавляет аромат, цвет, текстуру и стабильность при хранении.Роль сахара в том, чтобы сделать нашу жизнь слаще, несомненна.
ССЫЛКИ
Как производятся продукты. «Сахар». http://www.madehow.com/Volume-1/Sugar.html.
Кооператив по производству свекловичного сахара Южной Миннесоты. «Сульфитация». https://www.smbsc.com/OurSugar/SugarProcess/Sulfitation.aspx.
«Сульфитация». https://www.smbsc.com/OurSugar/SugarProcess/Sulfitation.aspx.
Сахарная ассоциация. «Знакомство с древнейшим подсластителем природы». http://www.sugar.org/sugar.
долларов США. 2020. Сахар: мировые рынки и торговля .Министерство сельского хозяйства США, Иностранная сельскохозяйственная служба.
История сахарной свеклы | CropWatch
Роберт М. ХарвсонОтносится к столовой свекле
Сахарная свекла, какой мы ее знаем сегодня, получена в результате многолетней селекции одомашненной свеклы ( Beta vulgaris L.). Говорят, что он получил свое название от греческой буквы бета , потому что раздутый, похожий на репу корень напоминает греческое B . Однако самый старый из известных видов свеклы, мангольд, был одомашнен как минимум в 2000 г. до н.э.C, и выращивался как греками, так и римлянами. Первоначально мангольд использовался в медицинских целях и из-за его плотного роста листвы в качестве горшечной травы, очень похожей на шпинат или некоторые китайские листовые овощи, которые используются сегодня.
Опухший, мясистый стержневой корень, известный сегодня, не был известен до второго и третьего веков нашей эры. Свекла, как красная, так и белая, была выведена в Италии (поэтому известная как «римская свекла») путем отбора из дикой свеклы, произрастающей на морском побережье. Средиземноморья. Он также был обнаружен по всей Европе и гибридизирован с сортами листовой свеклы (мангольд), чтобы получить широкий спектр цветов и форм, характерных для столовой свеклы сегодня.Его использовали как овощ, варили в рагу, запекали в пирогах и жарили целиком. Белая свекла, по-видимому, была более распространена, но менее желательна, чем красная.
В 18 веке крупнокорневую свеклу, известную как мангель-вурцель, скармливали скоту. Они были завезены в Англию в 1770-х годах для использования в качестве корма для скота после того, как были выведены из ранней кормовой свеклы в Германии и Голландии. Неудачный перевод немецкого mangold-wurzel («свекла») на английский как mangel-wurzel («корень дефицита») привел к убеждению, что это растение будет отличной пищей для бедняков в периоды голода, но оказалось, лучше подходит для коров.
свеклы были завезены в Северную Америку американскими колонистами, но когда точно неизвестно. Они были хорошо известны к восемнадцатому веку, поскольку упоминается мангольд, а также красная, белая и желтая свекла, выращиваемые в садах США в начале 1800-х годов. Джордж Вашингтон проводил эксперименты с ними в Маунт-Вернон, и к 1888 году Берпи Farm Annual предлагал семь различных видов мангелей, двенадцать разновидностей столовой свеклы и один вид мангольда.
Разработка современной сахарной свеклы
В середине 1700-х годов немецкий химик Андреас Марграф обнаружил, что и белая, и красная свекла содержат сахарозу, неотличимую от сахарозы, получаемой из тростника.Он предсказал тогда, что домашнее использование и производство сахара возможны в умеренном климате, но эти идеи не будут реализованы в течение следующих 50 лет, пока не будут разработаны новые способы добычи.
Один из учеников Марграфа, Франц Карл Ахард, проводил исследования в этой области, и благодаря его успеху в превращении свеклы в экономический источник сахарозы в Европе его теперь считают отцом сахарной свекловичной промышленности. Он построил первый сахарный завод в Кунерне в Нижней Силезии (современная Польша) и разработал эффективные методы переработки, используя плохие источники зародышевой плазмы из генофонда белого корма
Он построил первый сахарный завод в Кунерне в Нижней Силезии (современная Польша) и разработал эффективные методы переработки, используя плохие источники зародышевой плазмы из генофонда белого корма
, доступного в то время.Развитие свеклосахарной промышленности Германии, Франции и других европейских стран можно проследить до этих скромных начинаний, поскольку он свободно делился результатами этих исследований с другими.
До этого сахар получали только из тропического сахарного тростника, и для большинства европейцев он был непомерно дорогим. В начале 1800-х годов большая часть сахара была получена из Вест-Индии. После того, как поставки были прекращены из-за английской блокады континентальной Европы во время наполеоновских войн, спрос на сахар вырос по всей Европе.Наполеон поощрял новые исследования сахарной свеклы, и между 1810 и 1815 годами было введено в эксплуатацию более 79 000 акров земли, и во Франции было построено более 300 небольших заводов. После падения Наполеона сахар снова стал доступен из тропиков, и цены на него упали из-за избыточных поставок. Большинство заводов было закрыто, и новые разработки шли медленно. Однако после упадка рабства в Вест-Индии европейская промышленность стала более конкурентоспособной по сравнению с тропическим источником сахарозы, и к 1850-м годам эта промышленность прочно утвердилась и работала на большей части континента.
Большинство заводов было закрыто, и новые разработки шли медленно. Однако после упадка рабства в Вест-Индии европейская промышленность стала более конкурентоспособной по сравнению с тропическим источником сахарозы, и к 1850-м годам эта промышленность прочно утвердилась и работала на большей части континента.
Создание свекольных заводов в США
После завоевания места в Европе были предприняты многочисленные попытки наладить производство сахарной свеклы в США. Первая попытка вырастить сахарную свеклу была предпринята в 1830 году в Филадельфии, но фабрика так и не была построена, и от этой идеи отказались. Первая фабрика, построенная в США, была построена в Нортгемптоне, штат Массачусетс, в 1838 году, но прекратила работу после 1840 года. Были предприняты и другие безуспешные попытки основать фабрики в Висконсине, Иллинойсе и Мичигане.
В начале 1850-х годов мормонские пионеры начали исследовать местное выращивание сахарной свеклы и переработку сахарозы в качестве домашней промышленности, чтобы стать более независимыми. В 1852 году они основали фабрику недалеко от Солт-Лейк-Сити из машин, привезенных из Англии и доставленных в Юту крытым фургоном из Канзаса. К сожалению, из-за отсутствия знаний о химической обработке эта попытка также не увенчалась успехом для производства кристаллизованной сахарозы, но она проложила путь для будущего развития выращивания и переработки сахарной свеклы на западе Соединенных Штатов.Первое успешное коммерческое производство свекловичного сахара в США началось в центральной Калифорнии в 1870 году. К 1890 году две фабрики работали в Альварадо (теперь известном как Юнион-Сити) и Уотсонвилле.
В 1852 году они основали фабрику недалеко от Солт-Лейк-Сити из машин, привезенных из Англии и доставленных в Юту крытым фургоном из Канзаса. К сожалению, из-за отсутствия знаний о химической обработке эта попытка также не увенчалась успехом для производства кристаллизованной сахарозы, но она проложила путь для будущего развития выращивания и переработки сахарной свеклы на западе Соединенных Штатов.Первое успешное коммерческое производство свекловичного сахара в США началось в центральной Калифорнии в 1870 году. К 1890 году две фабрики работали в Альварадо (теперь известном как Юнион-Сити) и Уотсонвилле.
На протяжении всей истории производства сахарной свеклы в США многие заводы были запущены, но работали лишь в течение короткого периода времени. Эти начальные усилия часто предпринимались методом проб и ошибок, часто перемещаясь с места на место, пытаясь найти то правильное сочетание факторов, которое привело бы к большему долгосрочному успеху.Многие из возникших проблем были связаны с импортом семян сахарной свеклы из Европы. Исследователи из Министерства сельского хозяйства рано узнали, что лучшие результаты были получены при использовании местных сортов, но промышленность продолжала настаивать на использовании импортных продуктов. Такой образ мышления продолжал препятствовать постоянному успеху производства свекловичного сахара в определенных областях.
Производство свеклы сегодня
Сахарная свекла, как она известна сегодня, уникальна среди пищевых растений, используемых во всем мире, поскольку она является продуктом селекционных исследований, а также стала образцом для улучшения продуктивности растений с помощью генетики и селекции.Завод по производству сахарной свеклы также уникален в истории своей ролью катализатора революции в сельском хозяйстве. Выращивание сахарной свеклы в Европе в то время заменило прежнюю мелкозерновую монокультуру и ввело понятие севооборота. Это создало систему для удобрения и повышения плодородия почвы и уменьшения проблем с сорняками, а также предоставило источник пищи для домашнего скота из ботвы, кроны и жома свеклы.